细胞质杂交
出处:按学科分类—生物科学 农业出版社《植物细胞培养手册》第149页(11001字)
细胞质杂交是植物细胞遗传学中的一种新方法,它可能与作物改良有重大关系,这个方法是将具有不同的质体和线粒体的二个体细胞的细胞质融合到另一个细胞的原生质膜中,形成含1一2个功能核的异质细胞,即胞质杂种细胞或杂种细胞。通过细胞器不同的原生质体间的融合或把异源细胞器引入一个植物原生质体中,我们即可得到这样的杂种细胞。我们感兴趣的是使杂种细胞的细胞分裂能够进行下去的那些系统,以及细胞器在细胞分裂和再生植株的动向和表达,和由杂种植株得到有性后代的试验。细胞质杂交对以后植物细胞器遗传学的影响,并指出细胞质杂交如何与其它育种步骤结合起来,取得作物改良的成功。
可通过各种杂交手段改良栽培种。通常,尤其在质量性状成为问题时,植物育种者知道他需改良哪些性状,并且手头有一个或几个栽培品种,这些栽培品种除了个别需改良的性状外是令人满意的。因此育种者面临的问题是将所需性状“插入”现有栽培品种中。如果需改良的性状受核基因控制,杂交和随后在自交或回交后代中的适当筛选,通过适当的基因分离可获得改良品种。雄配子发育和其它单倍体化手段可缩短通常长达几代的筛选期。如果在栽培品种中没有所需性状,可采用种间杂交,但这些杂交可能会遇到各种不协调的障碍。在一些情况下,配子产生和胚开始发育,但进一步的发育停止。在这种情况下,应采用体外胚培养。然而栽培品种所需改良的性状受细胞器控制时,这种杂交手段并不很适用。如前面讲到的,杂种通常只含母本细胞器;因此细胞器控制的性状不会进一步分离。但也存在明显的例外(如燕麦),这在特殊作物中应考虑到。另一方面,在体细胞杂交中,我们期望首次融合产物含有融合双亲细胞器:这样的细胞称为异质细胞。这方面的证据由Gleba(1979)提供:他融合叶绿体控制的白化病烟草植株的原生质体与核控制的叶绿体缺乏的烟草原生质体,得到具有混合叶绿体的后代植株,并且在体细胞杂交的有性后代中发现杂色植株,这表明融合产物的异质特性。下面将提到另一个表明融合物中细胞器混合的例子。这样,目前已充分肯定体细胞杂交是一可能的获得新的核基因组/细胞质基因组组合的手段。另一方面,体细胞杂交也造成一些麻烦。有时我们只对叶绿体和线粒体基因组的变化感兴趣,而对核基因组的变化无兴趣。在这种情况下,我们应采用细胞质杂交,获得含非杂交核和异胞质基因的融合产物。这种异胞质基因组和同源核的融合产物,通过细胞器的分开和再分化,应产生所期望的植株。Cocking(1982)指出不同细胞器基因,如细胞质雄性不育基因,经原生质体融合后,在体细胞杂种细胞第一次有丝分裂时,观察到不同细胞器基因的单向分配。
(一)体细胞杂交中细胞器和胞质基因组性状
被子植物体细胞杂交中细胞器和胞质基因组性状的相关性怎样?有几种回答。第一,由胞质基因组控制的形态学性状和生化特性可确定体细胞杂交种。第二,某些性状如链霉素抗性和叶绿体缺乏可作为在群体或植株中筛选异源融合产物的手段,这些群体和植株由非融合和自融的原生质体形成。第三,为育种目的或遗传学研究,我们可能对现有栽培品种的线粒体和叶绿体的交换感兴趣,或对具有另一植株细胞器的基因型感兴趣。表6-11归纳了主要有关体细胞杂种植株中叶绿体、线粒体和雄性不育的信息。这儿将讨论几个特殊例子。有关通过胞质杂交传递细胞器的方面将加以详细讨论。
1.通过胞质基因性状确定体细胞杂种 由表6-11可见,只有有限数量的叶绿体性质用于确定体细胞杂种植株的叶绿体基因组。其中大部分如fentoxin反应、RUDPC酶大亚单位的等电聚焦Lp DNA的限制性模式和链霉素抗性已由Galum(1982)详述。具正常叶绿体的原生质体与具(叶绿体基因组控制的)缺陷叶绿体的原生质体融合,可产生第二种原生质核基因型的绿色或杂色体细胞杂种。这样,绿色或杂色在一些研究中被用作异质叶绿体基因组状态的叶绿体标记。目前唯一的线粒体标记是mt DNA的限制性模式:在研究适当例子时,将讨论这种标记以及细胞质雄性不育(CMS)的利用。
被子植物体细胞杂交中最先报道的是蓝格斯多夫氏烟草和粉蓝烟草的体细胞杂交。陈等(1977)分析了16个体细胞杂交植株的HUBPC酶。所有这些植株都具有双亲RUPBC酶的小亚单位,表明是细胞核的杂交。16株中,9株只具蓝格斯多夫氏烟草叶绿体大亚单位,而6株只具粉蓝烟草叶绿体的大亚单位;一株同时具有二种大亚单位,但它们在有性后代中分离。这些结果清楚表明融合伴随着叶绿体离析而产生异质叶绿体基因组状态,在多数情况下,叶绿体分出有些突然。大多数已报导的细胞质杂交都普遍存在着迅速离析。
等电聚焦常用作叶绿体标记(表6-11),因在一个试验中即可确定叶绿体基因组,另一方面它缺乏敏感性。因此使用这个方法Iwai等(1981)确定在普通烟草+黄花烟草的体细胞杂种中只存在普通烟草RUBPC酶的大亚单位,但通过这个杂交种的花药培养得到的一些植株,具黄花烟草RUBPC酶的大亚单位,这个体细胞杂种可能同时含有普通烟草和黄花烟草叶绿体,但后者在等电聚焦中低于测定水平。
表6-11 体细胞杂种植株的叶绿体、线粒体和雄性不育的分析

另一常用的叶绿体标记是cpDNA限制性模式。它首次由Belliard等(1978)用来分析普通烟草和迪勃纳氏烟草叶绿体。由于不同种的cpDNA可由一些核酸内切限制酶切成不同的片断,这种方法作为体细胞杂交中叶绿体的标记是相当有用的。另外通过各自限制性模式的分析,可测出cpDNA的种内差异。Scoweroft和Larkin(1981)用这种方法研究两个迪勃纳氏烟草变异体原生质体融合后的叶绿体的离析。
在一些植物如烟草、玉米、小麦和矮牵牛属中,CMS是严格由母体传递的。有很好的证据表明CMS在烟草属中同时受核和线粒体控制;CMS因此可起线粒体标记的作用。Belliard等(1978)检出二个烟草系间的体细胞杂种。一个烟草系是CMS(具迪勃纳氏烟草细胞质)和具有无柄叶(核标记),而另一个是可育的(具普通烟草细胞质)和具有柄叶。这两个系的有性杂种的叶型是中间类型。因此可区别体细胞杂种和胞质杂种。叶形或雄性不育/可育与任一亲本不同的植株被假设为体细胞杂种或胞质杂种。叶绿体DNA限制性分析曾用于确定试验植株的叶绿体,但未发现叶绿体类型和CMS间的联系。然而mtDNA限制型与CMS有关,最终,雄性不育被Ichar及其同事用来确定两个矮牵牛种融合后植株中的体细胞杂种。
mtDNA限制性模式只在两项研究中帮助确定体细胞杂种(或胞质杂种)。一项将在下一节中详细讨论。另一项对普通烟草和奈特氏烟草间体细胞杂种的研究由Nagg等(1981)完成。在这项研究中,在6个杂交无性系中分析了mtDNA限制性类型。除了大部分片段与双亲的相同外,这些杂种还具有独特的片段。基于考察各自的限制性类型与E.coli.DNA,提出在杂种mtDNA中发生了顺序重排。由于烟草种mtDNA限制性类型相当稳定,即使在离体细胞培养的情况下,重排应归于原生质融合后的异质胞质基因组状态。
2.利用胞质基因组性状筛选体细胞杂种 因被子植物中缺乏适合的线粒体突变株,下述讨论将限于叶绿体基因组性状。实践中用了两种类型的叶绿体突变株。第一种丧失光合功能,如缺乏叶绿素。第二种类型抗抗生素(如链霉素,卡那霉素),它们作用于叶绿体中的翻译过程,但不影响胞液中的翻译过程。链霉素抗性被Malige及同事用于一些体细胞杂交的研究中,用于筛选杂交植株(见表6-1)。例如,Medgyesy等(1980)将抗链霉素普通烟草系的原生质与林烟草原生质体融合,并将得到的愈伤组织移入含1mg/ml链霉素的介质中。普通烟草原生质已经碘醋酸盐预处理,因此若未与林烟草原生质体融合,它们不能存活。未融合和自融合的林烟草原生质体融合,它们不能存活。未融合和自融合的林烟草原生质体可预期产生对链霉素敏感的愈伤组织。这样,链霉素存在下再生的植株是体细胞杂种或胞质杂种。结果证实了这种预见。
表6-1 用细胞表现型的互补或部分互补找出体细胞杂种细胞或植株a

a.NR=硝酸还原酶;5MT=5甲基色氨酸;AEC=氨乙基半胱氨酸。
白化症和其它叶绿体缺陷症可由胞质基因组或核基因组引起。Gleba及同事利用这个性状筛选体细胞杂种或胞质杂种。融合双亲都为普通烟草原生质体,但一个具叶绿体基因突变(P-),而另一个是“硫”突变株(Su/Su),它导致黄色或致死植株(除非在含糖介质中培养)。绿色或绿、黄杂色植株可被筛选为融合产物。
(二)细胞质杂交方法
为了对获得具某种核基因组但具有外源线粒体和叶绿体的植株,我们应避免常规体细胞杂交即杂合核基因组带来的麻烦。在这方面,具有全套核、线粒体和叶绿体的原生质体与缺核或核分裂受阻遏的原生质体融合是有利的。另一方法是将叶绿体、线粒体或它们的DNA引入正常原生质体中,随后培养和植株再生。
这里只介绍融合和植株通过“供体受体”法再生的过程(图6-4)。
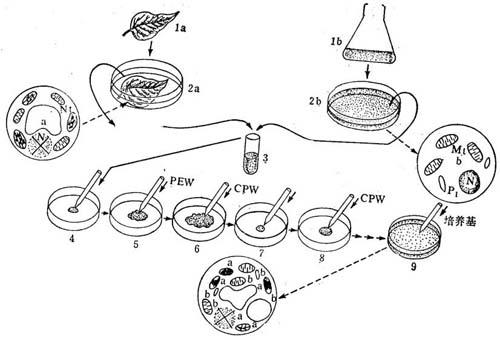
图6-4 胞质杂交的供受体方法的程序图解
la=供体叶;1b=细胞悬浮物(受体);
2a=供体叶浸入酶液中;2b=受体细胞悬浮物浸在酶液中;
3=混合X-辐照的受体和未照射的受体原生质体;
4=把0.25ml混合原生质体悬浮液插入培养皿的中央;
5=加0.35mlPEG溶液;
6=加0.5mlCPW;
7=除去溶液,剩下凝集的原生质体;
8=加0.5ml CPW;
9=最后用液体培养基稀释融合的原生质体;供体(a)和受体(b)原生质体含有核(N),线粒体和叶绿体或质体,融合产物原生质体仅只-个功能核。
1.原生质体
①选择“供体”和“受体”。二者在核和细胞质性质上都应完全不同。所需细胞器性质应在“供体”中。
②由细胞悬浮体或叶肉或二者制备原生质体。“供体”和“受体”原生质体来自不同组织时有好处,因能观察到异源融合和估计其百分比。在原生质体的离析和融合中应避免强光。
③X-射线照射“供体”原生质体。烟草属叶肉原生质体应照射的适当剂量是5Kγ;其它原生质体可能需要不同剂量以阻止核分裂。可用γ代替X-射线。
2.融合融合操作程序由Kao和Michayluk法(1974)修改而成。
①“供体”和“受体”原生质体按1∶1混合,最终密度各约为每毫升5×105个。
②用直径为6cm的塑料培养皿(组织培养级,如Falcon3002)并加入0.25ml的原生质体悬浮体于每个培养皿的中心。每个试验用几个这样的培养皿。
③以小滴形式加0.5ml PEG溶液于每个叶绿体悬浮滴的外缘。静置15min。
④以小滴形式加0.5ml CPW溶液于每个叶绿体悬浮滴的外缘,静置10min(可见原生质体聚集于小滴中心)。
⑤用巴斯德吸移管小心从小滴上去掉溶液(而不是原生质体,)并按上法另加0.5ml CPW溶液,置10分钟并重复这个步骤二次。
⑥加1-3ml液体培养基于每个培养皿中,用倒置显微镜观察确定特定原生质系的原生质体密度(烟草的最适原生质体密度为2×104个/m1),使用适合的特定原生质体的培养基。
⑦用石蜡膜密封培养皿并培养(通常25-28℃)。一或二天的暗或弱光培养可能对大多数原生质体有好处。随后暴露在50-2001x的光下。
3.生长和筛选
①2-3天后加入琼脂培养基,使琼脂最终浓度为0.8%(将42℃的琼脂培养基从小滴加入以避免原生质体突然加热)。在加入琼脂培养基前观察到有效原生质体密度过低于最适密度,将融合的原生质体转移到饲喂层上。
②除去未融合的和自融合的“受体”原生质体,可在液体培养初期进行(如以甘露醇为渗压剂,以除去林烟草原生质体),也可推迟到培养后期(如将幼小愈伤组织移入含链霉素介质中以除去对链霉素敏感的细胞)。采纳的筛选过程应适用于使用的特殊系统。
4.植株再生这里不详细介绍植株再生,由于诱发植株再生的步骤视各系统而异。
5.溶液制备
①PEG溶液 制备含50%聚乙二醇1500(如BDH Chemicals Itd,England)溶液,其中含10mM CaCl2和0.1M葡萄糖。
②CPW其中含0.55M甘露醇(最好来自BDH)、0.19mM KH2PO4;0.01M CaCl2,0.98mM MgSO4·7H2O,0.98mM KNO3,0.99mM kI和0.16mM CuSO4,溶于1000ml水中。
6.“供体-受体”方法——雄性不育的传递和细胞器交换 因“供体-受体”法有应用于一些系统的潜力,为此将其中有代表性例子进行适当的介绍。
Evans等第一个“供体-受体”试验的基本原理如下:未经处理和具已知胞质基因组的经X-光照射过“受体”原生质体,和具有不同胞质基因组的“供体”原生质体融合,随后在避免“受体”原生质体分裂的条件下培养,应该形成具“受体”核和“供体”胞质基因组性状的胞质杂交植株。我们选林烟草为“受体”,是因为它的原生质体在含甘露醇的介质中不分裂。从cms烟草系(L-92)得到“供体”原生质体,据报道它是由香甜烟草(雌)与普通烟草(雄)杂交和随后不断与普通烟草为授粉者回交得到的。L-92的形态学与普通烟草Xanthi品种的相同;但它为cms,具类柱头/类花瓣花药和无生活力花粉。因是单亲传递,L-92叶绿体和线粒体相同或非常类似于香甜烟草的叶绿体和线粒体(因为不能确定香甜烟草或一个有关的原产于澳大利亚的烟草种,是否事实上是L-92的原母性祖先,将L-92的细胞器称为“L-92”而不称为“sua”)。分别用核基因组、胞质基因组和雄性不育(或可育)的缩写形式表示,第一个融合试验可写为:
syl/tbc/fert+tbc/L-92/ster(irr.)这意思是:林烟草核/普通烟草叶绿体/雄性可育十普通烟草核/L-92叶绿体/雄性不育;irr.表示X-照射。普通烟草和林烟草叶绿体的所有性质相同(如tentoxin抗性、经一些限制性内切酶降解后cp DNA模式型),因此用tbc表示这二个种的叶绿体。
期望异源融合的原生质体具有筛选上的优点。因在他们的标准(甘露醇)培养基中,X-辐照过的“供体”原生质体不能生长,“受体”原生质体不能分裂,而后者只有与“抗甘露醇”的普通烟草原生质体融合后才能分裂。结果可归纳如下:大多数再生植株(来自5个愈伤组织的20个植株)具林烟草的形态学特征,但和“供体”植株一样雄性不育。这20株被称为类型-A植物。通过使用三种叶绿体标记(tentoxin敏感性、RUBPC酶的等电聚焦和cp PNA的限制性类型)发现类型-A植株有L-92叶绿体。因此这些叶绿体和CMS通过CMS烟草系(L-92)传递到林烟草。类型-A植株,就叶绿体成分和CMS的一致性,经用林烟草和普通烟草异交以及雄核发育而得到证实。因此,类型-A植株可表示为syl/L-92/stero
这些结果使我们提出如下问题:(a)与可育“供体”融合是否能恢复类型-A植株的可育性?(b)可育性的恢复是否与叶绿体的转移有关?
第一个问题的肯定回答表明“供体-受体”方法普遍可用于单向转移雄性不育和雄性可育。另外,如果叶绿体的转移与雄性不育无关,后一性状可能受线粒体控制,结果“供体一受体”方法的应用更为广泛:叶绿体基因组的性状可与线粒体的性状独立地转移。
为回答这些问题,又进行了如下融合:
syl/L-92/ster+tbc/tbc/fert(irr)即,以类型-A为“受体”,和以正常普通烟草Xanthi为“供体”。在一个试验中,我们得到63个愈伤组织,其中13个肯定为融合产物。这些愈伤组织产生以下杂种植株:
7株来自3个愈伤组织:syl/tbc/ster
19株来自8个愈伤组织:syl/tbc/fert
8株来自4个愈伤组织:syl/L-92/fert
在一些情况下,愈伤组织产生二种植株(如,syl/tbc/fert和syl/L-92/fert)。另外有一株,syl/L-92/ster具有与类型-A核/叶绿体/不育性相同组成的植株,很明显它是融合产物,因为产生它的愈伤组织同时也产生syl/tbc/ster植株。很明显在愈伤组织水平上质体的离析仍在进行。很少观察到一个融合产物愈伤组织分化出雄性不育和可育的植株。这可能意味着线粒体的离析比叶绿体的离析要早得多?还没有回答这个问题,但应指出细胞构架(cytcskeleton)可能提供胞液中线粒体间的相互联系。如果被子植物中的实际情况就是如此,一个细胞的线粒体组成,可能不能认为是由一定细胞内的物理上独立的粒子所组成。
这些试验进一步提出如下问题:(a)在融合产物的细胞分裂中线粒体离析吗?(b)线粒体基因组与雄性不育/可育相关吗?(c)在异质胞质基因组融合的胞质杂种中,叶绿体与线粒体独立分配吗?我们可通过融合双方(如类型-A和普通烟草cv xanthi)的mt DNA限制性模式的明显不同来回答这些问题。还有两个观察值得注意。普通烟草和林烟草mtDNA的限制性类型无区别(在用xhoI或SalI切割后)。另外,细胞悬浮体和完整植株的mt DNA限制性模式相同。在每个胞质杂交植株中,mt DNA的限制性模式是稳定的:即使一定胞质杂种的有性后代也保持原胞质杂种的类型。分析胞质杂种mt DNA得出的二个主要结果是(1)大多数胞质杂种具一些双亲所没有的片断;(2)雄性不育胞质杂种总具有与不育亲本类似或相同的mt DNA限制性类型,而雄性可育胞质杂种具有与可育系本身类似或几乎相同的mt DNA限制类型。
因此,这些胞质杂种可分组如下:

(*表示与相应的mt DNA类似或相等。)
后一发现表明线粒体组成与雄性不育/可育的相关性。这些发现与Belliard等(1979)和Nagy等(1981)的发现一致。mt DNA在有性繁殖和细胞分裂中是稳定的,与此相反,新的mt DNA的限制性模式表明在原生质体融合后mt DNA的碱基顺序发生了变化。这变化由重排或重组引起还有待研究确定。这里要指出,如果被子植物细胞中的全部或部分线粒体相互有联结,这为在一个胞质杂种细胞内,提供了不同线粒体基因组相遇的机会;这是一个对胞质基因组难得或不存在的机会,因为尚无迹象表明被子植物细胞中存在叶绿体的相互联结。
虽然,发现了雄性不育/可育与线粒体类型的相关性,对叶绿体则不存在这种相关性:常有这样的现象,具相同叶绿体组成的二个胞质杂种在育性上表现不同,而具有不同叶绿体组成的胞质杂种在雄性育性上表现相同。
前述的三个问题是这样回答的:线粒体的离析独立于叶绿体的分配,并且线粒体的分配与雄性不育/可育的分离相关。
上述试验和其它类似的“供体-受体”试验在Evans实验室中进行了论证。表6-12表明,尽管从6个试验中得到的愈伤组织的数量不同,这些试验却产生了大量胞质杂种愈伤组织。另外,从每个试验中都获得大量胞质杂种植株。链霉素抗性似乎是一个有效的筛选方法,它只导致少量的“漏掉”(“escapee”)愈伤组织。抗链霉素性从普通烟草SR-1“供体”转移给林烟草。应指出,在这个特定试验中大多数胞质杂种只产生抗链霉素的后代,而一些杂种同时产生敏感的和抗链霉素的苗。表6-12中试验5和试验6的设计是为了产生杂色植株。这样的植株存在于相应的杂种中,表明叶绿体的完全离析并不一定是细胞质杂交的必然结果。
表6-12 6个不同融合试验得到的胞质杂交植株
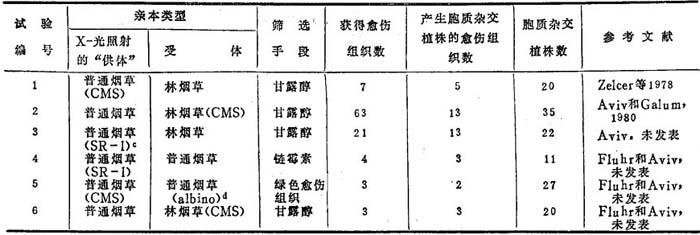
a.选取“受体”(“供体”原生质经X-辐射后停止分裂)。
b.胞质杂种的鉴定,除基于至少一个胞质标记外,还基于形态学特征.
c.链霉素抗性突变体来自Dr.P.Maliga,Szegea,Hungarg。
d.白化苗衍生于一具缺陷叶绿体的杂色的普通烟草。
7.修改的“供体-受体”法使用吲哚乙酸
Maliga及同事对“供体-受体”方法作了值得注意的修改。这些作者用由Wright(1978)提出的吲哚乙酸失活技术来筛选融合的哺乳动物细胞。这项技术以经吲哚乙酸预处理的原生质体代谢被抑制为基础。这种预处理使未融合和自融合的原生质体退化,而预处理的原生质体与未处理的原生质体融合致使代谢上的补偿并产生有生存力杂种。在这一试验中,吲哚乙酸预处理过的蓝茉莉叶烟草细胞悬浮体原生质体与X辐射过的普通烟草叶肉原生质体融合,得到47个愈伤组织。所有来自这些愈伤组织的(胞质杂种)再生植株具有蓝茉莉叶烟草的形态学特征,但它们中的大多数含普通烟草的叶绿体。吲哚乙酸处理没有损伤原生质体的核,因此处理过的原生质体可补偿X-辐射的原生质体,因此可预期形成胞质杂种。以类似方法使用其它代谢抑制剂也是可行的。
【参考文献】:
〔1〕Carlson,P.S.,Smith,H.H.and Dearing,R.D.1972 Parasexual interspecific plant hybridization.Proc.Nat.Acad.Sci.69:2292-2294
〔2〕Evans,D.A.,Wetter,L.R.,and Gamborg,O.L.1980 Somatic hybrid plant of N.glauca and N.tabacum Obtained by protoplast fusion.Physiol.Plant.48∶ 225-230
〔3〕Kao,K.N.and Michayluk,M.R.1974 A methed for high-frequency intergeneric fusion of plant protoplasts.Planta 115:355-367
〔4〕Kao.K.N.1975 Nutritional requirements for growth of vicia hajastana cells and protoplasts at a very low population density in liquid media.Planta 126: 105-110.
〔5〕Aviv,D.and Galun,E.1980 Restoration of fertility in Cytoplasmic male sterile(CMS)Nicotiana Sylvestris by fusion with x-irradiated N.tabacum protoplasts.Theor.Appl.Genet.58:121-127