折返的基本机理
出处:按学科分类—医药、卫生 军事医学科学出版社《临床心电图学词典第二版》第262页(1866字)
形成折返的基本条件为:一个有效的折返环路、单向阻滞和足够缓慢的传导。
显然,折返性心律失常的决定因素是传导障碍。作为折返基础的传导障碍有下列类型:(1)快反应动作电位转变为慢反应动作电位,产生缓慢传导和单向阻滞:心肌细胞的静息膜电位水平,是决定传导速度的因素之一。静息膜电位水平愈负,则有更多的钠通道被激活,钠离子进入细胞内的速度也愈快,这就是快反应动作电位。静息膜电位在-80~-90mV的心肌纤维及浦氏纤维都能发生快反应动作电位,其传导速度约1~4m/s。
当急性心肌缺血或梗死时,局部心肌的严重缺血可诱发持续的局灶性电活动,使静息膜电位降低。类似的情况也见于各种情况造成的心肌损伤中。当静息膜电位在-60~-70mV时,仅有50%的钠通道被激活,此时动作电位O相的峰值速度和振幅均低于正常,传导速度也减慢。这是由于静息膜电位较低,因而钠离子内流进入细胞较慢所致。
此称受抑制的快反应动作电位。当静息膜电位低于-55或-60mV时,动作电位主要是由钙离子并伴少量钠离子内流引起。
当静息膜电位接近-50mV时,钠通道失活,钙通道开放,产生慢反应动作电位。在人的病变心脏,既能产生受抑制的快反应动作电位,亦能引起慢反应动作电位。
激动在心肌纤维中呈双向传导。如快反应动作电位受抑制,激动在两个方向的传导呈同等程度减轻,单向阻滞并不出现。
但如产生慢反应动作电位,则激动在一个方向呈缓慢传导,在另一个方向完全受阻,便形成单向阻滞。(2)激动的相加法则:激动相加(summation)法则是由Cranefield在1971年提出的。激动的相加机制是形成单向阻滞的又一原因。激动的相加首先需要传导纤维的一种特殊排列,即两支纤维汇集到一支纤维的解剖结构。
从两支纤维传来的激动如同时抵达汇集处,可相加成一个较强的激动,通过汇合纤维。如该汇合纤维的某一节段组织是受抑制的,则激动能否通过此抑制区,就取决于激动的强度。
如激动能相加成一个较强的激动,就能通过该区;如-个激动提前抵达而不能和另一激动相加,则这个提前抵达的激动就不能通过该抑制区;如激动是从逆向通过此区则是激动的分散而不是相加,结果造成阻滞。可见在这种情况下冲动的相加可以形成单向阻滞,如不能相加则此抑制区是双向阻滞。
心脏的许多部位都具有这种心肌纤维汇合的解剖结构,在一定条件下即可发生激动的相加和单向阻滞。(3)激动的抑制:激动的抑制(inhibition)是描述两个传播激动互相作用的又一种情况,即当一个激动本身不能通过某抑制节段,但却首先抵达该抑制区,则使该节段处于不应期,这样从另一个纤维束传来的原先能通过该抑制区的强激动亦受到阻滞。
由于一个较弱的激动的首先抵达使较强的激动受到阻滞就是激动的抑制,因而也可以是单向阻滞的一个原因。激动的抑制同样可以发生在有纤维汇合的部位,由于能形成单向阻滞所以同样是形成折返的一个可能的机制。(4)激动的反折:折返的另一种形式是通过激动在传导途径中反折(reflection)引起的,反折是使激动能在抑制节段返回的又一种机理。反折是指在两条紧邻的抑制纤维,如一条受抑制较深,则激动在此纤维受到阻滞,而在另一条抑制较浅的纤维内缓慢传导,如传导足够缓慢则激动波峰就能扩散到较深抑制的纤维内而返回,并在深抑制区缓慢传导。
如正常心肌组织已脱离了有效不应期,则激动将又一次激动正常心肌组织形成一次折返激动或折返性心动过速。反折引起折返的特点是并不需要环形的解剖结构,即可形成激动的返回。这种反折较多见于心室内的浦氏纤维束中,如心肌梗死区周围心肌组织中就可能形成反折。此外,沿心肌纤维不同走向所形成的*各向异性折返,也在折返激动的形成中占有重要地位。参见“各向异性结构”。
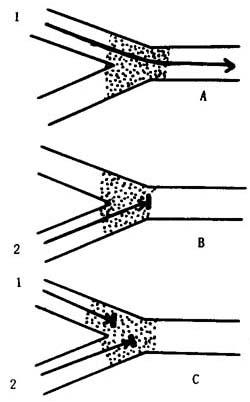
激动的抑制
A.较强的激动能通过抑制区。
B.较弱的激动不能通过抑制区。
C.较弱的激动首先抵达,使较强的激动也不能通过抑制区。
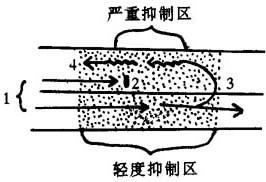
激动的反折机理
图示两条邻近的浦氏纤维,上条抑制较深,下条抑制较浅,激动(1)将在较深抑制区被阻滞(2)而在另一条中慢传导,激动波又经深抑制区返回(3)(4)。如此时正常心肌组织已脱离了有效不应期,则形成一个折返激动。