青藏高原鼠疫自然疫源地研究进展
出处:按学科分类—自然科学总论 北京出版社《现代科技综述大辞典上》第1388页(3907字)
青藏高原鼠疫自然疫源地(简称疫源地),是世界海拔最高、中国面积最大而发生人类鼠疫较频繁的地区之一。
鉴于该疫源地发现较晚,可供借鉴的历史资料缺乏,而鼠疫动物病的流行又较猛烈,为此对其进行深入研究,不仅对揭示鼠疫菌、宿主和媒介三者在一定生态地理环境中的生物学变化及相互依赖关系,鼠疫动物病的流行范围和年际与季节变化规律颇有裨益,无疑也将有助于阐明该疫源地性质、结构乃至保存规律的认识,从而为控制、监测和消灭鼠疫提供科学依据。
该区位于欧亚大陆东南部,介于东经79°51′~103°20,和北纬30°40′~39°21′之间。
北起祁连山,南止喜马拉雅山,东临甘南草原,西与克什米尔相望。地势西高东低,境内气候干寒多风,温湿度和降水量由于各地水热状况不同差异很大。植被自东南向西北由半湿润的高山草甸草原过渡到高山寒漠或半干旱高山草原。此地区啮齿动物具有种类较少而数量较多的特点,并以高原特有种占优势。
其中,喜马拉雅旱獭(简称旱獭)、多种鼠兔、高原鼢鼠等分布广而数量多。
自1954年首次在青海省贵南县上网察(今贵德)与河南蒙旗克萨木地区分别自旱獭和达乌尔鼠兔分离出鼠疫菌后,相继于1959、1973、1976年发现甘肃、新疆和西藏旱獭疫源地。
近40年来,在该疫源地分离鼠疫菌近2000千株,判定疫源县(镇)40余个,疫源面积20余万平方公里。证实啮齿动物旱獭、达乌尔鼠兔、五趾跳鼠、灰仓鼠、根田鼠、小家鼠和食肉兽艾鼬、狗獾、兔狲、猫、猞猁、沙狐、赤狐、牧犬,以及有蹄类绵羊、藏原羚与猛禽胡兀鹫等17种动物自然感染鼠疫(兔狲、沙狐、胡兀鹫以血清学判定);斧形盖蚤、谢氏山蚤、腹窦纤蚤深广亚种、红羊新蚤、人蚤、具钩靴片蚤、圆指额蚤上位亚种、草原硬蜱、血红扇头蜱和旱獭虱等10种吸血节肢动物自然携带鼠疫菌。其中,旱獭为主要宿主,斧形盖蚤和谢氏山蚤为主要媒介。旱獭系青藏高原特有种和优势种,分布广,几与该区高山草甸草原和高山草原相一致,并具有连续分布特征。
垂直分布在海拔1900(循化孟达)~5450m(冈底斯山)之间,其中以2800~3600m低中山下部或丘陵平原、河谷阶地数量较多,调查资料表明,每公顷两只以上的土地面积占总面积的2%,1~2只的占15%,不足1只的占83%。但据1989、1990年青海、甘肃、西藏调查发现,每公顷仅为0.22~0.50只,显着低于60~70年代水平。
旱獭为草食性动物,以禾本科的、莎草科的和豆科的植物叶、茎为食,在田边亦啮食庄稼,有时也吃甲虫。
在饲养条件下,除配方饲料外,还吃白菜、甘蓝和葫萝卜等。
为穴居冬眠动物,每年10月初入蛰,4月初出蛰。3龄性成熟后每年繁殖一次,约50%~70%的雌獭参加繁殖;胎仔数2~6只。
旱獭以家族洞系为中心,活动半径一般不超过200m,当有强烈干扰时可做2100~5120m较远距离迁移。旱獭洞分居住洞和非居住洞两类。居住洞又分永久居住洞(冬眠洞)和季节居住洞(夏洞)两种;非居住洞又分取食避难洞和废弃洞两种。永久居住洞巢室在0.08~0.23m3之间。冬眠时旱獭体温维持在5~7℃(室内1~2℃),略高于环境温度2~3℃(室内0~-2℃)。旱獭呈吻尾相对球形。
其呼吸、心跳分别为18次/min和28次/min;取出洞外5min分别增至22~28次和52~64次,20~30s后剪趾始见血液渗出。
据青海、甘肃、西藏1982~1991年统计,旱獭自然检菌率波动在0.22%~0.39%之间,占总检菌数的98.08%。实验感染表明,初次感染10亿菌可全部死亡,再感染700亿菌尚有存活个体,可见再感染旱獭对鼠疫菌有较高耐受性(武文莲,1982)。
用捕自疫区的和非疫区的旱獭实验,发现两区旱獭死亡率无显着差异(t:0.82 P≥0.05),但疫区旱獭完全致死量为50亿菌,非疫区旱獭50亿、5亿、50万均可完全致死。
前者死亡时间集中在感染后的3~8d,后者在9d内。表明两区旱獭对鼠疫菌均具较高感受性和敏感性,但前者个体差异大,有一定抗性,感染后可呈迁延带菌过程;后者则较差(朱锦沁,1983)。
旱獭血凝阳性检出率为8.47%;感染后旱獭血清都具有相当保护力(MPI<10)。其抗体在感染后5d出现,15~30d滴度最高,两个月后可见转阴个体,冬眠时滴度下降,13个月后滴度基本消失。
自本区发现蚤类160余种,斧形盖蚤和谢氏山蚤为旱獭鼠疫主要媒介,占獭体蚤90%~100%。但随气候和植被变化,高山草甸草原以斧形盖蚤为主,有向高山寒漠草原或半干旱高山草原以谢氏山蚤为主过渡趋势。
旱獭染蚤率在42%~72%之间,蚤指数为1.17~4.87。每年5、9月各有1个数量高峰,前者以斧形盖蚤为主,后者以谢氏山蚤为主。
研究发现,两蚤均为1年1个世代,分4个龄期。斧形盖蚤♀∶♂1∶0.56,春末为吸血、产卵和数量盛期;谢氏山蚤♀∶♂1∶0.49,秋季为吸血、产卵和数量盛期。
并发现,吸过血的蚤受精囊内基本都充满精子;新羽化的斧形盖蚤(未吸血)可忍饥180余天。
据青海1982~1991年统计,斧形盖蚤和谢氏山蚤自然检菌率分别为0.11%~0.36%和0.07%~0.21%,前者占昆虫总检菌数的44.68%,后者占42.55%。
媒介效能研究正在进行中。
经对阿胶糖、鼠李糖、麦芽糖、密二糖、甘油和脱氮等生化试验,可将该区鼠疫菌分为青藏高原型的,型连山型的,阿尔金山型的,昆仑山A、B型的和冈底斯山型6个生物型。其毒力因子均具有F1+、Pstl+和约98%及86%左右为VW+、Pgm+菌株,对小白鼠最低致死量在10~10000个菌。Pgm+突变Pgm速率高于85%。
自该区鼠疫菌检测到6、7、13、23、27、30、45、52、62、65和90MD11种质粒。
其中大质粒分布区,52MD主要见于祁连山及青海湖周围,且为青藏高原所特有;65MD主要见于阿尔金山及青南高原;90MD主要见于西茂安多、那曲和青海小唐古拉山地区。
在免疫学研究中,已选育出FI+、VW+、Pgm和Psti+0614F为新菌苗株。ELISA抑制法检测血清抗体,具有用一种酶标记物检测不同种属动物的可能性。
经化学试剂诱导,本区已建立FI-;Pgm-;VW-;FI-、Pgm-和FI-、WV-、Pgm-和5种毒力缺失型菌株,为进一步研究疫源地提供了基础资料。
青藏高原为以旱獭为主的单宿主疫源地。动物病多发生在山麓或冲积平原凸丘阳坡或半阳坡;疫源性活跃而牢固,动物病频发,多呈散在流行;以6~8月为季节流行高峰,新的疫源地区和染疫动物不断被发现。根据该区鼠疫菌生化特性(方喜业,1980)和自然屏障(王振华等,1990)各分10个生物型和独立疫源地。
鉴于该区人、鼠间鼠疫流行态势,现已实施的省、州、县三级监测网仍是今后控制鼠疫的主要技术措施。
鼠疫菌保存机理是当今世界性研究课题,现仍限于各种假说而无定论。
本区根据旱獭巢微小生物群落中的线虫与旱獭蚤的寄生关系,提出鼠疫菌在该环境中可能存在线虫 蚤
蚤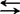 宿主双项循环关系(詹心如,1989)。而更多的人则依据宿主(媒介)对鼠疫菌具有一定选择作用和鼠疫菌在通过不同环境时具有菌株变异特性而进行的一系列研究。如鼠疫菌质粒种类的研究和建立毒力缺失型菌株等,都将为揭开鼠疫长存之谜创造条件。鼠疫监测也是现今控制鼠疫的主要手段和研究内容。青藏区实施的政府与专业组织相结合的三级监测网是行之有效的,但以间接监测和以微机管理取代直接监测将为期不远。
宿主双项循环关系(詹心如,1989)。而更多的人则依据宿主(媒介)对鼠疫菌具有一定选择作用和鼠疫菌在通过不同环境时具有菌株变异特性而进行的一系列研究。如鼠疫菌质粒种类的研究和建立毒力缺失型菌株等,都将为揭开鼠疫长存之谜创造条件。鼠疫监测也是现今控制鼠疫的主要手段和研究内容。青藏区实施的政府与专业组织相结合的三级监测网是行之有效的,但以间接监测和以微机管理取代直接监测将为期不远。
【参考文献】:
1 武文莲,等.中华流行病学杂志(鼠疫论文专辑),1985,30
2 纪树立.鼠疫.北京:人民卫生出版社,1988.44~316
3 王淑纯,等.鼠疫研究进展.北京:中国环境出版社,1988.1~305
4 朱锦沁,等.鼠疫研究进展.北京:中国环境科学出版社,1988.369~374
5 詹心如,等.鼠疫研究进展.北京:中国环境科学出版社,1988.280~287
6 王文华,等.鼠疫研究进展.北京:中国环境科学出版社,1988.346~350
7 王振华,等.鼠疫研究进展.北京:中国环境科学出版社,1988.207~217
8 纪树立,等.中华流行病学杂志,1990,(特1):29~32
9 方喜业.中国鼠疫自然疫源地.北京:人民卫生出版社,1990.34~71
10 李少春等,中国地方病防治杂志,1990;5(增刊):107~109
(青海省地方病防治研究所李少春副主任医师撰)