再生植株的遗传变异性
出处:按学科分类—生物科学 农业出版社《植物细胞培养手册》第171页(8762字)
(一)产生变异的技术
采用组织培养循环产生植物有6种不同技术。这6种产生体细胞无性系变异的特殊操作技术是:(a)长期培养循环;(b)原生质体培养循环;(c)愈伤组织培养循环;(d)特殊组织外植体的利用;(e)伴随特定营养基或激素配比选择导致随机变异的产生;(f)某种导致变异数目增加的基因型的利用。
事实上,在这个过程中所用的外植体可取自植物体的任何组织,包括叶片,节间,子房,根和花序。正是由于这些类别范围极广,因此他们相互联系。例如,长期组织培养物或特殊外植体来源的利用必定要结合原生质体培养或愈伤组织培养一起来做。
表8-1是对23种植物产生变异的情况以及获得这类变异所采用的技术的概括。在大多数情况下,只对那些由组织培养直接获得的植株及其营养繁殖体作了观察。至少已有10篇报道证实这种特性可有性传递(表8-1)。所有这些情况都说明变异可稳定地传递到后代。
表8-1 孢子体组织培养体再生植株中已证实有的变异
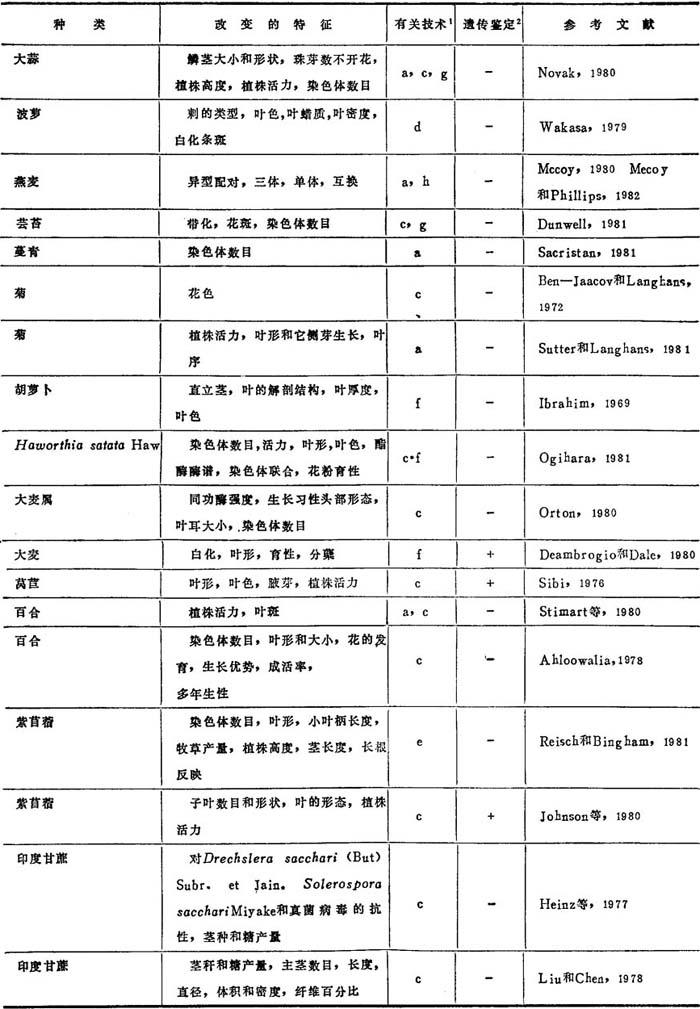
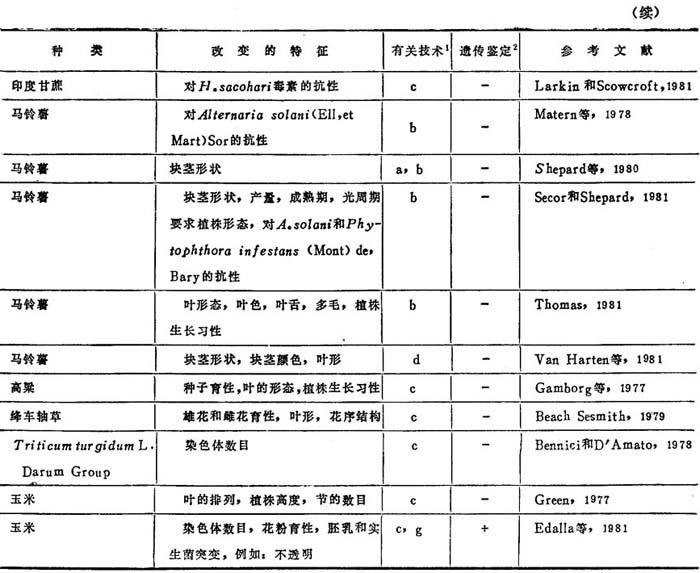
1.a=长期培养循环;b=原生体体培养循环;c=愈伤组织培养循环;d=特殊组织外植体的应用;e=伴随组织培养抗性选择的随机变异世代;f=特殊营养培养茎或激素组合的利用;g=倾向于变异数目增加的某种遗传型的利用。
2.+=所改变的特征已确证可有性传递;一=所改变的特征来观察到可有性传递。
1.长期培养循环长期培养物再生植株的可能性随着种类的增加正在变成现实,这在过去常是很困难的,有关长期培养再生植株的形态和遗传稳定性已有少量报道。现已知道,随着培养时间的增加将产生核型结构的变化;因此,植株不正常性的频率很可能随培养时间的增加而增加。但是,一般情况下,再生植株的变异性并不反映培养细胞中所存在的整个不正常性范围,这表明在再植株前发生了一些选择。
经长期继代培养的烟草愈伤组织再生植株,其花和叶的不正常性很普遍。但在相同培养条件下,这种不正常性在短期培养物再生植株或由种子培育的植株并未发现。然而,已注意到这种不正常类型的愈伤组织可能已在长期培养过程中无意之中和有选择性地得到了保存。
在菊花嵌合植株中曾观察到短期茎尖培养物再生植株只有少数不正常性,而由另一维持9年之久的栽培品种叶愈伤组织再生的植株都是高度变异的。这些变异包括:植株矮化,叶形改变,花变小且形状不规则,侧芽过度生长,这同只培养一个月的叶愈伤组织再生植株相比显得非常明显。虽然,嵌合重排是一种可能性,但也不能排除残留激素的影响以及遗传不稳定性这类可能性的存在。
McCoy(1980)证实,在燕麦栽培品种Lodi和Tippecanoe中,细胞遗传上的不正常植板的频率随培养年龄的增加而增加,幼年培养体中产生的不正常植枝经常只有简单的改变,而有20个月年龄的培养体再生的不正常植株,经常出现一条染色体改变。相反玉米的长期培养物都是稳定的,对培养8个月的愈伤组织再生植株的分析,表明有94%的植板是正常的。
2.原生质体培养循环 首先,原生质体培养再生植株在形态染色体数目以及育性方面都发生改变。但是,在其它有关原生质体再生植株的一些报道中,常可观察到表型的稳定性。由四种烟草种原生质体获得的植株,在表型上没有改变。营养体和花的形态以及染色体数目,在所有再生植株中都是正常的。但这个结论只是根据20株植株观察得到的结果。
Wenzel等(1979)在马铃薯原生质体培养获得的植株中也观察到这种明显的一致性。有些植株虽然在块茎形状上发生了变化,但这些变化仅仅在延长培养期后才出现。所观察到的少量变异,可归于培养过程中诱发的非整倍性所致。这个结果与Shepard等(1980)利用栽培品种Russet Burbank获得的结果不符。Shepard筛选出由这个种原生质体获得的一万个无性系,结果发现有大量园艺上的重要变异。其中有些无性系的块茎形状、产量、植株形态以及对早、晚疫病的抗性都产生了变异。这种变异的遗传基础一直未得到确定,但对这些无性系中的65个无性系作了详细的分析,采用分析的性状有35种,包括:叶,块茎,花以及整个植株的特性。在这项研究中采用的65个无性系是由1700个原始群体中挑选出来的。剔除严重的不正常类型,所选的无性系在外表上都比较正常并且具有相当良好的生活力,蔓型以及块茎均匀一致。这65个无性系中每个无性系至少在某一特性方面与亲本Russet Burbank不同,有三个无性系只有一个特性发生变化。有一个无性系却有17个特性与亲本不同。
Shepard和Wenzel的工作中存有这种不一致性可有几种解释。首先,Wenzel等(1979)所用的马铃薯品系与Russet Burbank不同。结果当然完全与Russet Burbank不同。早已证实,在培养过程中不同的遗传型可产生不同数目和类型的变异,此外,Wenzel仅筛选少量(211)无性系,而Shepard筛选了10000个以上。另外,在采用的技术方面也有一些差异,包括:(1)原生质体的来源(Shepard用叶肉;Wenzel用茎尖培养物);(2)倍数性水平(Shepard用4x;Wenzel用2x)。有趣的是Thomas(1981)报道,在由四倍体British马铃薯栽培品种Maris Bard茎尖培养物获得的原生质体再生植株中,观察到大量的变异。这些变异包括:生长习性的变化,叶形,颜色,光泽度以及多毛性状。从不同的原生质体获得的25组植物中,仅仅只有2组与亲本栽培品种Maris Bard相类似。因此,变异显然可由茎尖培养的原生质体或叶肉原生质体再生植株中产生,但至今为止,在双单倍体马铃薯无性系中还未观察到存有体细胞无性系变异。
3.愈伤组织培养循环 通过组织培养途径创造遗传变异,最广泛采用的技术是愈伤组织培养循环。经愈伤组织培养再生植株具有高度变异,这一点已由Barbier和Dulieu(1980),Sibi(1976),Edallo等(1980),Dono(1978)以及其他工作者(表8-1)作了很好的记载。Sibi(1976)发现,由莴苣愈伤组织培养物再生植株,在形态上发生改变,这些再生植株自交的子代在叶的特征,腋芽发育以及叶色上都发生了变化。某种叶的特性显然是细胞质发生变化的结果,因为这些特性均为母系遗传。将三种交异株与对照株进行双列杂交,结果表明,异型杂交比自交子代显示更强的活力,即便是在原始材料纯合的情况下也是如此。其它一些变化,在进行三次自交时也从不分离,由此推测也是母系遗传的。但这并不排除由于染色体结构变化造成育纯突变体的可能性。
相反,Edallo等人(1981)从玉米二个自交系未成熟胚获得的组织培养物再生植株中发现,仅有核DNA发生变化。通过遗传研究,已证明大多数变异体是明显可见的基因突变。McCoy(1980)早已报道,在细胞遗传上,玉米显然是稳定的。Edallo等(1981)发现,110个再生植株中只有二株与二倍体植株不同,而没有观察到染色体内变化,总的来说,每个植株大致显示一种单基因突变。
Barbier和Dulieu(1980)利用核基因来追踪直接由(1)子叶,(2)子叶获得的原生质体,(3)不同时间间隔的愈伤组织,诱导形成的烟草植株中存在的植株变异模式。缺失和倒位可说明所观察到的这种高频率的叶色变异。作者通过对三种不同组织培养物处理所得到的变异进行比较后认为,“再生使上次细胞循环后,累积在子叶静止细胞中潜在的变异性得以表达;这种变异性也可在培养细胞中累积”。
在由800个水稻愈伤组织获得的体细胞无性系中,所有观察的性状有28.1%具有正常的遗传模式。通过二次自交对这些改变的性状进行分析的结果表明,这些植株在叶绿素含量,花期,植株高度,育性和形态方面均发生了改变。由此认为,这些变异或是已存在的整体变异或未分化细胞生长期间积累的变异。
4.特殊的植物组织培养 在相同的条件下,采用的外植体不同可明显地影响变异的性质和类型。在凤梨中,由四种不同类型外植体(果皮,接枝,冠瘿,腋芽)得到的愈伤组织再生植株,其特性随外植体来源不同而变化,由接枝,冠瘿和腋芽得到的植株中只有刺的特征发生改变,而由果皮得到的一些变异在叶色,刺,蜡质和叶密度方面都发生变化。由此可得出结论,只要通过对外植体适当的选择,就可采用组织培养方法来迅速繁殖无变异的小植株,或诱导产生有用的变异。Van Harten等(1981)的工作表明,只要仔细选择外植体就有可能增加体细胞无性系变异体的产生。由小叶叶缘产生的植株,有12.3%在表型上发生了变化,而由主轴和叶柄获得的愈伤组织产生的植株,有50.3%在表型上发生了变化。
5.伴有生化细胞选择的变异 前面详细的讨论体细胞选择方式。由于在规定的培养基上可相当容易地培养大量的植物细胞,因此曾设想这种系统有微生物型细胞操作的优点。最初,有关这方面的工作一直受到经选择过的细胞无能力再生植株的限制。最近,这类选择的细胞可再生植株,并对再生植株作了观察,而且还对这些再生植株的子代所选特性的遗传稳定性作了核对。若干由变异培养物得到的植株中,除那些正在探索的一些特性外,其它特性均发生了改变,这表明在所选变异体细胞内存在有简单变化或多重变化的基因多效性效应。
这种体细胞无性系变异类型,也许与采用细胞选择系统之前,通常经诱变处理培养物产生的变异是不同的。在组织培养条件下,常不能确定培养物产生的变异是诱变发生的,还是随机地经过与有关体细胞变异的其它一些结果更一致的过程产生的。Berlyn(1980)通过对烟草单倍体细胞培养物进行辐射处理,选择到抗异烟酸酰肼(INH)的变异细胞系。这些细胞培养物再生的植株,在生活力、根的发育,叶形和生长习性上都发生改变。其中缓慢生长的特性与INH的抗性有关,通过杂种与亲本进行回交可加以分开,这表明这些特性是独立遗传的。
在含有乙硫氨酸培养基上,经选择培养的二倍体苜蓿培养物可再生植株,当将这些植株与原始植株进行比较时发现,在有些性状上发生改变。93个细胞系中,有23个再生的植株,在染色体数目、叶形、小叶柄长度,牧草产量、植株高度、茎长度以及发根的情况都发生了改变,其中有些特性的改变对苜蓿的育种可能是重要的。因此,这一结果表明农艺上所要求的无性系可通过组织培养循环得到改良。
在这项研究中,尽管所观察到的大多数变异体是在含有乙硫氨酸的条件下选择培养的,但对乙硫氨酸的毒性并不具有抗性。这说明在许多细胞系中均带有抗性和非抗性的细胞。同时也说明所观察到的广泛的变异类型对乙硫氨酸的抗性都是完全独立的。同样,在这项研究中还在植板于无抑制剂的培养基上再生了一株四倍体植株(NSI)。这一植株显然比对照的四倍体细胞系产生更多的牧草干物质,这表明它在其它特性上同样存在差异。从苜蓿愈伤组织再生植株中通常变异频率很低。显然,个别的变异体虽可由苜蓿组织培养体中产生,但结合在乙硫氨酸条件下培育来选择时,得到的变异频率更高。
Reisch和Binghan(1981)指出,在乙硫氨酸或EMS和乙硫氨酸共同存在的条件下,再生植株均可发生改变。经EMS诱变处理,但未植板在乙硫氨酸条件下的对照组中,再生植株中只有极少数(61株中有3株)发生改变。事实上,乙硫氨酸可能是诱变真核生物细胞的诱变剂。用乙硫氨酸处理大豆种子和玉米颖果的结果表明,即便单独使用乙硫氨酸也可产生诱变。因此需通过进一步试验来确证,经乙硫氨酸处理的培养细胞,在用或不用诱变剂处理时,是否能同样诱导有用的变异。
生长培养基和激素的效应。Bayliss(1980)对有关培养基和激素,尤其是2,4-D对细胞培养体的影响作了很好的描述。但是在再生植株过程中有关培养基成分影响的详细报道不多。当将大麦组织培养体放在含有4.5-18.0μM2,4-D的条件下4-56天,这些培养体仍可再生植株,对这些再生植株的后代进行观察结果,发现存在着遗传变异。这些变异包括白化、叶形和分蘖的育性,而这些变异的特性只有在18.0μM2,4-D的条件下才产生。
Haworthia愈伤组织培养物再生植株有各种不同的细胞遗传特征,曾对这些植株的细胞遗传学变化作了详细观察,用于再生植株的二种培养基仅在激素含量上存在差异。这二种培养基是:NK培养基(26.8μM NAA和0.46μM KIN)和I培养基(0.57μMIAA)。在NK培养基上再生的植株要比I培养基上再生的植株,具更多的四倍体植株,而带有染色体易位的植株则很少。
遗传型反应。在仔细控制培养操作和再生顺序的情况下,再生植株中的变异性可能是独立遗传的。Lin和Chen(1976)对8个甘蔗栽培品种试验结果发现,在所观察的4600株植株中,大多数植株与其供体存有差异;F146号无性系形态变化的频率是1.8%,而F156号无性系为34.0%。
Skirvin和Janick(1976)对Polargonium sp栽培品种愈伤组织培养再生植株的形态学变化作了详细观察,结果发现变异类型的产生取决于最初所用的栽培品种。因此,他们指出,这种变异性对无性系内,尤其是对具多倍性的有性繁殖的无性系内植株的改良可能是有用的。
(二)体细胞无性系操作在植物育种中的作用
长期以来,植物育种者一直认为,有选择地改良一个普遍的栽培品种,远比创造一种新品种更有价值。培育者往往更愿意种植较老的栽培品种,因为这种栽培品种在局部地区已被证实是有价值的,并对其生长特性已为培育者所熟悉。虽然体细胞无性系操作可改良植株的性能,但新的栽培品种未得到检验,还不了解他们的消费市场。因此,目前用来增强普遍栽培品种性质,而又保存其基本个性的技术基本上有5种:(1)回交育种;(2)无性系的选择或芽变选择;(3)突变育种;(4)组织培养体中产生的变异体,以及(5)单基因转化。纯合的自花授精作物可通过上述任何一种方法来完成。然而,在一种较理想的栽培品种中,仅缺少一种或二种简单的遗传特性,因此,回交育种就是通常采用的选择方法。在杂合的营养繁殖作物中,采用回交育种的方法就不能重新创造优良的基因组合,因此也就不能保存轮回亲本的杂合性质,即便仅用于改良单个无性系也要遇到这个问题。为了有选择地改良这种作物就必需利用其它的方法。
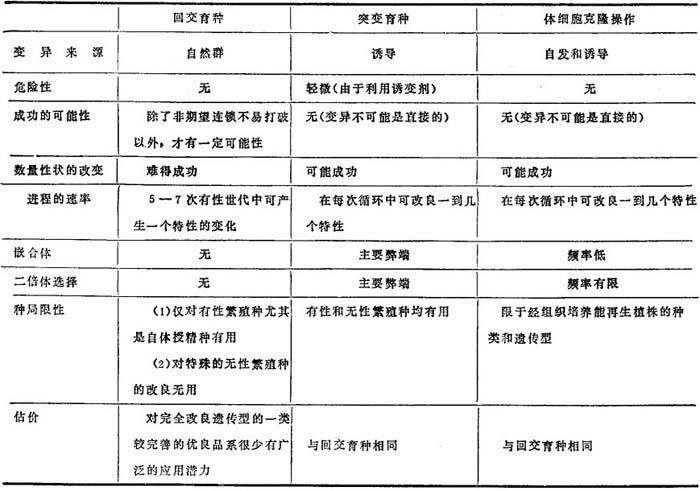
其它选择方法在应用上也是有限的。至今为止,高等植物单基因转化的方法,还未能足以发展成为一种有用的工具。偶然的芽变选择,需要大规模的种植、敏锐的观察以及增强这一难得的事件产生的机会。在喷洒农药种植的情况下,就不可能发现抗病虫害的芽变,此时突变育种就是一种可供选择的方法,并可结合体细胞无性操作新技术一起加以应用。表8-2对突变育种技术与体细胞无性系操作以及回交育种的方法作了比较。
表8-2 普遍栽培品种或育种品系产生单一变化所采用的三种方法的比较
(三)体细胞无性系变异是突变育种的一种形式
实际上,经体细胞无性系变异进行突变育种和植物改良是相同的二项技术。体细胞无性系变异可揭示原先就存在于完整植株中的遗传变化,或发生在培养的细胞中的变化。在组织培养循环中,经体细胞无性系变异发生的变化,在逻辑上可考虑为突变育种过程的延伸。这个过程可包括涉及到遗传变异的任何一个过程。
有关马铃薯突变育种研究的结果表明,离体培养技术可作为诱导变异的一种方法,以替代诱变处理。在马铃薯突变育种中曾采用离体条件下诱发不定芽技术,并对组织培养技术产生突变诱导和嵌合现象的效能作了观察。营养组织突变发生嵌合现象的问题,是一个十分复杂的问题。由不定芽产生的植株一般是不是嵌合的。但在这项研究中,当对离体培养产生的不定芽用γ-射线处理时,获得的植株却出现极高的突变频率,包括极广阔的突变系列和极低的嵌合率。意外的是,由叶轴和叶柄未经辐射的对照中(不是叶外植体)获得的植株具50.3%的突变频率。这个频率几乎与辐射系列一样高。在早先用相同栽培品种所做的一些试验中,诱导突变的频率只有24-38%,因此,人们曾设想“在马铃薯突变育种中不用射线处理,从而可避免辐射诱变处理产生的这种不需要的边隙效应”。显然,这些研究者并不知道,原生质体和悬浮培养物再生植株中存在有广泛变异的文献,但却分别得出了利用组织培养再生植株中发生的变异来改良作物这一概念。
(四)当前的应用
对于体细胞无性系变异体的研究使一些有价值的育种品系得以建立。目前,在台湾试验场栽培有从甘蔗愈伤组织培养物获得的高产抗黑粉病的品系。有关甘蔗中其它细胞无性系变异体的工作也取得了进展,产生了一些具重要的潜在变异能力的无性系,这些无性系的大部分性状与其亲本相同,但具有抗霜霉病和斐济病的特性,而这是一种流传广又很重要的病毒。已建立有若干抗斐济病的无性系,这些无性系不具有明显有害的不正常性,在所有出现的性状中,只有一个性状与原始的无性系Pinder差不多,其它性状如茎杆产量和蔗糖百分含量与原始无性系有些差异。这些重要的性状对于证实选择优良的性质是必需的。如同其它任何一种技术一样,新并入到普遍栽培品种去的特征,对于测定时空遗传型的反应仍是必需的。体细胞无性系产生变异的技术,仅对特定遗传背景内产生的新的变异有益,但对于最终的测定还必需利用常规的育种手段。以及对栽培品种的相应优良性能作出估价和加以选择。
Skirvin和Janick(1976)对天竺葵属中产生的体细胞无性系变异进行深入研究后,建立了一种具香味特性的天竺葵,他们称为Velvet Rose。可以相信这是通过组织培养技术建立的第一个得到命名的栽培品种。
体细胞无性系变异在增强遗传物质的交换方面也可能是有用的。Larkin和Scowcroft(1981)对广泛的有性杂种中有用的外来基因的渗入作了描述。黑麦草和大麦杂种胚培养产生的植株是高度变异的,其农艺性状要比那些不通过组织培养循环产生的有性杂种要有价值。同时,在杂种中由于作物和外来基因组间缺少交换而阻碍了外来基因整合到作物植株中。杂种胚培养的结果表明,细胞培养显然打破了只有在生殖细胞中才能进行遗传物质交换的界限,杂种胚的愈伤组织状态可克服这一障碍。
【参考文献】:
〔1〕Larkin,P.J.and Scowcroft,W.R.1981 Somaclonal Variation-A novel souree of variability from cell Cultures for plant improvement.Theor.Appl.Genet.60:197-214。
〔2〕Skirvin,R.M.1978 Natural and induced variation in tissue Culures.Euphytica27:241-266.
〔3〕Sharma,A.K.and Sbarma,A.1980 Chromosome Technigues.Theory and Practice,3rd ed Butterworths.London,Boston,Sydney,Wellington,Durban,and Toronto.