葡萄
出处:按学科分类—生物科学 农业出版社《植物细胞培养手册》第572页(20405字)
葡萄属(Vitis)是Vitaceae科(前称Ampelidaceae)的唯一属,有商品价值。根据化石记录,早于人类历史很久。小亚细亚是其栽种发源地。公元前400年向西传至埃及和北非,公元前2000年传入中国。
葡萄(Vitis vinifera)有上千个变异体,是唯一商品种。经开拓者、殖民者和定居者带至南和北半球。有时,偶见的葡萄与本地野生种异种间杂交产生有价值栽培品种,Concord葡萄是其一。
葡萄属是温带植物,不能忍受高纬度和高海拔地区的极低温度。夏季光充足是最适成熟所需。目前分布在北纬50°到南纬40°之间,海拔高度1500-2500m。法国300m以下地区质量最好,中等质量的可产于约1000m地带。
葡萄组织培养与常规育种有机配合,有可能产生佳味特性,抗冬季低温,土壤和种子携带的病害和虫害的栽培品种。
(一)研究进展
Morel(1944)首次报道葡萄培养成功。测定了细胞增殖的最适生长素浓度,建立了不依赖荷尔蒙的细胞系,和需较低浓度生长素的细胞系。也指出了茎外植体的愈伤组织的起源。Kulescha(1949)证实了驯化的冠瘿细胞(也是驯化的)比需生长素细胞生产生长素量更大。
50年代大多数人工培养研究,集中于解剖学的,化学的和酶的差异和培养的葡萄冠瘿细胞的维生素要求。Slabecka-Szweykowski(1952)报道了光、糖类和渗透性亏缺对驯化的细胞内花青素产生的调节作用,Fallot(1955)指出了建立几个种和栽培品种的细胞培养体试验。Duhamet(1951)发现了椰乳增强细胞生长及其与生长素的互作,Hilderbrandc(1958)建立了葡萄phylloxera肿瘿的单细胞无性系。把需生长素、不要生长素和冠瘿细胞与幼蔓嫁接,证实了需生长素和驯化的细胞生长受寄主的控制,但肿瘤细胞不是这样。
60年代着重系统测定从phylloxera肿瘿分离的单细胞无性系和正常河岸葡萄组织需要的生长因子,昆虫肿瘿与正常克隆间生理和生化差异和它们对γ射线的不同敏感性。Reinert(1963)在规定培养基上建成了冠瘿细胞的单细胞克隆(以前取得的无性系生长于附加椰乳或其它复杂有机物的培养基上),Reinert(1964)培养了整体浆果,有些产生了愈伤组织。
最近研究最适微繁殖方法中,发现亲本蔓光周期处理能影响外植体的潜在形态建成。
60年代最有实践价值的工作,是用热处理产生无病毒克隆的方法,以及热处理茎叶外植体的生长和器官再生问题。
70年代离体培养葡萄研究,在差异性和数量方面大量增加,并开展了许多新的激动的工作。用数学分析了细胞和核大小间生理学相关后,结束了phylloxera肿瘿和正常细胞研究。增加了消除病毒研究,主要工作放在寻找热处理茎尖的生长和发育最适培养基化学和物理参数上。也描述了把热处理植株茎端帽嫁接在幼苗或草本根砧木上的方法。
研究了蔓上各种外植体如浆果和胚乳和葡萄栽培品种的四和二倍体无性系产生细胞培养体方法。研究了细胞的长期培养成功的条件和培养基成分。
Benbadis与Baumann(1973)首次报道葡萄原生质体培养,指出其细胞分裂和壁再生可能有困难。可是,果皮愈伤组织原生质体产生再生壁和愈伤组织,提出并非不可能取得成功。
花药培养产生了单倍体愈伤组织,继而死亡,再生植株和休眠类胚结构也未能生存,以及愈伤组织产生了有生活力的二倍体基因组体细胞胚。控制葡萄植株再生研究取得突出成效。葡萄和其它葡萄属未受精胚株,葡萄属杂种的未成熟茎,花簇和幼叶取得了体细胞胚。1977年春,在Montbray葡萄园,Silver Run,Maryland首次建成了种植由体细胞胚产生的蔓。
70年代期间,开发了葡萄快速无性繁殖的三种方法。Pool与Powell(1975)与Ecevit(1979)报道了用节间插枝繁殖所需条件,Jona与Webb(1978)与Harris与Stevenson(1979)报导了茎尖快速繁殖和消除病毒所需条件。Barlass与Skene(1978)叙述了茎尖分生组织切段外植体诱导产生的不定芽快速繁殖新方法。
广泛研究了离体葡萄茎叶的发育解剖和形态建成的调节。证实了离体培养下在有限程度上发生卷鬚形成、花和果实。在适宜荷尔蒙条件下,原基(一组分生组织细胞)能产生卷鬚或花原基,对了解促进花器形成有重要意义。体细胞胚,不定芽或反复继代的茎叶切段,产生了具有幼年特性的蔓。
为了解决离体葡萄细胞长期培养和更广泛栽培品种和种再生植株方法,研究了外植体年龄和形态学上的部位和氢离子浓度和光质与光强对葡萄细胞生长与分化的影响。
Grenan(1980)和Valat等(1981)报道了离体培养和热处理对继后形态学改变和“治癒(Cured)”根和接种的大田表象的影响。
Jona(1980)和Meredith(1981)讨论了用组织培养改进葡萄的展望和策略,并在应用组织培养于改良作物方面存在着巨大机会。
80年代,采用茎尖或茎叶切段繁殖法,使葡萄繁殖商品化。不断改进和最适合的不定芽的起源和产生茎叶的化学和物理参数。茎尖切段产生的茎叶曾用于选择和适应含盐逆境。
采用胚珠培养系统,使正常无籽葡萄栽培品种产生种子,将有助于育种规划,和增进了解胚的发育。
最近,胚珠培养产生体细胞胚,扩大到若干其它栽培品和种。论述了从其它体细胞胚产生的体细胞胚的调节有关的化学和物理因素。
可以预期针对细胞和离体茎叶培养体的抗逆性如盐、硼、pierce′s病毒素和低温,研究的巨大增加。寄主与病原菌共培养技术研究有所改进。其目的在于选择耐真菌病原菌(粉霉病)和线虫的克隆,和测定寄主一病原菌专化性的生化、生理和遗传性质。热处理方法改进,用不定芽繁殖或茎尖繁殖,可以配合运用,提供快速繁殖和病毒指数植株储存的商品生产过程。通过人工培养,从成年转变成幼年状态的性质,也应研究。也要开发用幼年组织作为全能性细胞的渊源。
(二)培养程序
1.建立细胞培养系统 采用葡萄细胞培养选择耐化学或物理性逆境存在二个主要问题:①某些种和栽培品种的外植体的细胞培养体难于建成和保持,和②建成的细胞培养体很少产生不定芽或胚。长期培养难点,有遗传和生理一生化基础。葡萄属和葡萄栽培品种的驯化的或冠瘿肿瘤细胞已取得长期培养物。不要荷尔蒙的系在培养中的寿命极为稳定。最近已取得了葡萄和其它葡萄种的长期培养体。关于取得长期培养成功的因素讨论如下。
(1)外植体源 植物和动物形成器官、组织和细胞需要基因组成的区分表达。在培养中,一定的离体外植体基因表达的稳定性,可以保持或消失,结果使得从同一植株但不同器官的细胞,对人工培养下生长要求的条件不同,并常产生表型不同的细胞。
幼茎、叶柄、卷鬚和芽外植体曾分离出相对稳定和生长快的细胞培养体,从叶肉,花序和果实细胞则产生了生长较慢的分离物。浆果,再生的果皮原生质体也取得了稳定游离体。Krul等从葡萄种cv.Seyval的体细胞胚取得了不要荷尔蒙细胞系。至今未报导过根外植体的培养物。细胞培养体的寿命,不象是外植体形态部位的函数;可是,这些游离体的基因型是主要决定者。
Hawker等(1973)测验了几种葡萄栽培品种浆果产生愈伤组织。除2个(Wortley Hall和Muscat Gordo Blanco)外,都产生了愈伤组织,其余8个产生中度到快速生长的愈伤组织。Staudt等(1972)指出葡萄栽培品种Riesting和Portugieser的四倍体无性系比其二倍体无性系产生的愈伤组织更多。由于每种倍数体水平的干重相同,可知四倍体无性系生长较大,是细胞体积较大(大部分是水)所致。
(2)季节影响 葡萄许多栽培品种是光周期敏感性,短日反应表现为生长率降低。分离物能在全部季节中产生培养体,但观察到生长率和形态潜势不同。例如,Kober5BB茎分离物,生长在短日下,主要产生根和少量愈伤组织,而在长日下生长的植株的相同外植体产生愈伤组织。Brezeanu等(1980)报道了几种葡萄栽培品种的愈伤组织快速启动、生长和形态变化的季节影响。观察了:二月份培养体表现长停滞期愈伤组织建成和最初几次继代生长慢,后来生长很快;四月和五月份培养体,愈伤组织经少数继代后产生了根,和十月份,愈伤组织易于生根。
(3)光 亲本植株所处日长,影响着愈伤组织启动速度和继后生长率。光对建成细胞培养体的效果很不稳定。Hawker等(1973)提出光对几个葡萄栽培品种浆果愈伤组织细胞生长并无好处。相反,Lai等(1980)证实了葡萄Sentenial愈伤组织生长与白色光强增高呈直线关系。Brezeanu等(1980)葡萄栽培品种愈伤组织生长,以光强5.6klx最适宜。
光质也能影响葡萄细胞生长。Lai与I(1980)证实了蓝光对愈伤组织生长的增强,比红、绿或黄光或黑暗相对较大。细胞是否从每种光谱接受了相等能量。
愈伤组织培养体虽然对光周期不敏感。虽然多数学者发现16h光照,生长充分,其他学者发现了相对短日12h较好。
(4)温度 葡萄细胞生长和发育对温度反应强烈。休眠芽和种子在恢复生长前均须冷冻处理。由此设想葡萄含有感温基因,它们调节着生长和发育。25-27℃能满足人工培养下细胞生长。phylloxera肿瘿细胞最适温度比正常细胞的(36-38℃)为低,提出对温度的要求,应寻求更多信息。
(5)H+浓度 葡萄Phylloxera肿瘿细胞生长最适pH为4.5,正常克隆的为pH5.0(Pelet等,1960)。当正常细胞用γ射线处理时,其生长的最适pH,比辐射的瘿瘤细胞的为低。葡萄Sentenial细胞生长的pH最适范围为5.2-5.6。这也适合于至今供试过的葡萄的大多数细胞培养系统。但是,河岸葡萄生长最适pH比葡萄为低,提出在启动耐Ca22+和感性栽培品种培养体时,培养基中的H+浓度应加考虑。
(6)营养盐类 曾在广泛矿质盐类组成的培养中建成了葡萄细胞培养体。无见优劣之分。
(7)碳源 葡萄细胞培养体生长以蔗糖58.4-87.6mM为宜。也能在葡萄糖和水解淀粉上生长。葡萄肿瘿细胞以淀粉水解物优于蔗糖。
Phylloxera叶肿瘿细胞在甘露糖中比正常细胞生长更快,在果糖上后者比前者生长为快。二类细胞对各种糖类最适浓度不同,肿瘤细胞的一个克隆的生长,乳糖与蔗糖效果相同。这项结果很有意义,由于乳糖对许多植物组织有毒性,并能调节形态建成活动。
(8)维生素 葡萄肿瘤细胞生长需要硫胺素,肿瘤细胞的其它克隆生长不需要维生素。泛酸钙能增强正常细胞生长,对Phylloxera肿瘤细胞无效,吡哆醇和氯化胆碱均无影响(Arya,1965)。长期培养体失败,是否与维生素缺乏有关。
(9)其它有关物质 葡萄细胞培养体生长并无需要,附加某些复杂有机物,有利于离体细胞的生长。椰乳(10%v/v),酵母提取液(10%w/v)(Pelet等,1960),硫酸腺嘌呤加水解酪蛋白(各为40mg/L和3-10g/L),水解酪蛋白(2克/升)和酵母RNA(10-41mg/L),均能加强离体细胞的生长率。
(10)生长调节剂 许多早期工作者采用了IAA,NAA,或IBA,加或不加椰乳,以诱导和保持离体细胞的生长。以后学者发现椰乳可用细胞分裂素代替。
苯氧乙酸盐(2,4-D,POPA或NOA)适用于愈伤组织启动,可能对体细胞胚诱导有重要性,但在其存在下,不是经常可能保持长期生长。
BA是用得最广泛的细胞分裂素,KIN或ZEA也有效。GA3,ABA或乙烯对葡萄细胞的启动和生长,未见报导。本文未能举出许多重要葡萄栽培品种的愈伤组织或细胞悬浮物的长期保持的技术。由于这种缺陷,利用葡萄细胞于新开发的作物改良策略(突变体,抗逆性选择,原生质体融合等等)受到严格限制。有可能某些栽培品种对培养基成分敏感,例如rhododendro”培养物对MS中“正常”钾水平极敏感或需要未经测试过的维生素或生长调节剂。系统研究培养基成分,可为保持长期培养体提供信息。
2.原生质体培养 采用原生质体以引入外源种质、体细胞杂交和研究荷尔蒙作用、膜结构和功能,和细胞壁合成,在典型植物系统中已知甚多。葡萄原生质体用作基因受体发展较迟。这是由于其原生质体再生尚待证实。体细胞融合未能产生葡萄杂种的原因有三:1.葡萄原生质体尚未能再生,2.尚未取得四倍体蔓和3.尚无单倍体植株。
从葡萄果皮愈伤组织、茎愈伤组织和冠瘿肿瘤细胞制取了原生质体。产生了细胞壁和分裂一或二次。至今,只有果皮的原生质体产生的愈伤组织有继续细胞分裂能力。
已发表的报道,都是用愈伤组织或液体细胞悬浮培养体分离原生质体的。以前者更为有效。
愈伤组织培养在26℃16h,用2%纤维素酶(Onozuka SS)和1%果胶酶(macerozyme)。最适渗透条件为保持在KCl-CaCl2溶液中(浓度分别为0.1和0.14M),pH调节到5.5。暗培中完成壁消化。当用系列酶在35℃消化细胞壁时,消化可加速,原生质体在少于2h内释放,但由此产生的原生质体,形成特殊的壁,和不能分裂。
原生质体纯化,细胞与酶混合液经系列不锈钢筛(网孔150和60μm)过滤,取原生质体100×g离心3min。然后用KCl-CaCl2洗3次,每次洗后,50×g离心3min,取得片状沉积物。
原生质体沉积物悬浮在适宜培养基中,调节细胞密度105原生质体/ml。取3ml原生质体悬浮液转移到6cm培养皿中,用石蜡膜封边。暗培26℃。暗培细胞成活百分较高,用琼脂培养基(6g/l)比液体悬浮体为好。
支持壁再生长和继续细胞分裂的培养基是Ohyama与Nitsch(1972)营养盐类和Gam-borg与Eveleigh(1968)的维生素,附加水解酪蛋白(2g/l),谷酰胺(13.7mM),蔗糖(29.2mM),核糖(1.67mM),葡萄糖(1.39mM),木糖(1.0mM),阿拉伯糖(1.0mM),甘氨酸(0.05mM),肌醇(5.5mM),丝氨酸(0.95mM)和BA(4.4μM)。
细胞壁再生培养基用2,4-D(0.45μM)和蔗糖(0.23M),细胞增殖用NAA(5.4μM)和山梨糖醇(0.24M)较快。如果培养基的渗透性超过0.5M,细胞不发育。将细胞克隆从壁再生培养基取出转移到Hawker等(1973)愈伤组织培养基中,在几次继代中继续分裂。
3.花药培养 如果有同质自交系,葡萄育种将可顺利得多。有性世代时间长和现有基因型的复杂性,自交(经2-3个世代后产生不育性)和选择有困难。几种作物育种规划中,已采用了秋水仙素处理花粉胚“立即”产生自交系。为了取得葡萄同质系,可能必需采用加倍单倍体,由于葡萄属尚未看到自然单倍体,和自交也只限于少数几代。
Greenhoff与Doy(1974)在27品种中只有3个的花药产生了单倍体愈伤组织。无一再生植株,但有少数形成绿色细胞,其中有维管束组织。3个月后全部培养体死亡。蘡薁(Vitis thumbergii)花药愈伤组织产生了再生植株和类似休眠胚的结构。再生植株未能生存。glore葡萄(葡萄×砂地葡萄)杂种的花药愈伤组织取得了体细胞胚和有生活力植株。方法如下。
(1)外植体预处理 长2-3mm具有单核期花粉粒为主的花药,经4℃处理72h后,接种培养。除去全部花丝,切取时注意不伤花药壁。
(2)诱导培养基 Nitsch与Nitsch(1969)盐类附加0.02M FeSO4·7H2O,1μMBA,和5μM 2,4-D,最适于产生愈伤组织,继而产生胚。附加NOA和/或较高浓度BA的培养基,产生的愈伤组织不能成活,或继代后停止生长。花药培养在装有25ml液体培养基的100ml欧氏三角瓶中,每瓶接种25个花药。放在摇摆培养箱上(摇摆80次/min),27℃。琼脂培养基不适于诱导过程。冷冻预处理和暗培是胚诱导所必要的。启动培养后10-20天,可看到突破花药的愈伤组织。
(3)表达培养基 生长在基础培养基或基础培养基附加GA3的愈伤组织或细胞团块,光培(16h照光,2.5W/m2)时,产生大量体细胞胚。启动后45天形成成熟胚。
(4)胚发育 胚在琼脂基础培养基上发育慢,只有2%的胚形成植株。GA3(1μM)增高正常植株发育频率(47%),胚在4℃冷冻处理2-8周,频率增高到83-94%。
冷冻处理对各个胚胎发生时期都有效。未经冷冻处理的胚产生的植株,第一三小叶叶在30天内出现;而处理的,在转到27℃后7天内形成。
总之,只有雄蔓或强趋雄性的产生胚。雌雄同株的Cabernet Sauvignon产生愈伤组织,最后褐化死亡。愈伤组织再生植株,全是二倍体(2n=38)。胚是否双单倍体或从体细胞组织而来还不清楚。产生了许多矮株和反常蔓。生存率和生长旺势有差异,只有25%植株移栽入土成活。
4.体细胞胚胎发生 采用现代遗传工程策略或从细胞大群体里选择耐逆性克隆取得成功的先决条件之一,是从离体细胞产生植株的技术学,后者也可用于新引种植物的集团繁殖,和/或保持和产生无病毒植物。用于从表皮细胞连续产生胚的稳定系统最新发展,和分离出具有胚胎发生竞争性的不要荷尔蒙葡萄细胞,可能为研究胚诱导和发育期间基因表达提供诸系统。
从离体薄壁细胞产生胚过程,包括三个可鉴别的阶段,以细胞形态学和对生长调节剂的反应为基础:(a)诱导或体细胞薄壁细胞转变成原胚胎发生的集团(PEM),(b)表达或PEM转变为胚,和(c)胚的发芽和发育成株。
(1)诱导 高度液泡化的薄壁细胞经内部分裂而转变成一簇小、厚壁、细胞质浓细胞,具有明显核和核仁,这种过程叫诱导过程。诱导细胞的确切部位尚属不知。可是,它们虽然从珠心callote产生的,和分离的愈伤组织表面细胞,体细胞胚的根和茎连接处的单个表皮细胞和胚柄。具有诱导的或可诱导的细胞的组织或器官有珠心callote,花药,叶柄和脉间区域,未成熟花簇和绿茎,胚柄和体细胞胚下胚轴与根连接处的表皮细胞。
亲本植物遗传组成对诱导过程有显着作用。葡萄栽培品种体细胞胚的诱导,限于珠心外植体,耐野生Vitis种和异种间杂种具有较大范围的可诱导组织或器官。
诱导过程的确切化学和物理需求尚待确定。设想诱导需要一种生长素。由于启动愈伤组织也要生长素,因而难于证实这种要求。如果启动的愈伤组织含有竞争力细胞,那末生长素作用可能有争论。但是,如果后来才产生竞争性,应考虑其它物质。
当用含有氧乙酸侧键生长素时,能更有效地以珠心组织诱导产生胚,也能诱导类胚结构和再生植株,但频率低,和再生植株发育受损。诱导也需要细胞分裂素,因为用基础培养基加BA培养的胚,转移到加BA加NOA培养基上的顺序处理,是诱导产生珠心体细胞胚的最有效途径。
葡萄Seyval表皮培养在无荷尔蒙培养基上,产生了胚。ZAA(0.1μM),GA(0.001-0.1μM)和蔗糖(0.2-0.4M)增高胚的产生。加活性碳(0.01-1.0%),乙醇(0.001-1.0%v/v),脱落酸(0.01-100μM),BA(0.01-100μM)或低温,胚诱导率降低。
用Nitsch与Nitsch盐类或MS盐类取得了体细胞胚。有一例,在Nitsch与Nitsch盐类而不是MS的盐类产生了再生植株和类胚结构。
诱导培养基的物理状态,是某些外植体的关键决定因素。胚珠外植体和花药产生胚以液体培养基比琼脂培养基为好,培养基的物理状态对从体细胞胚产生胚无恒常影响。
光抑制从花药产生胚,降低叶愈伤组织产生的二极类胚结构数在16h光周期下,从很幼年体细胞胚的胚的发育,比8h光照的为强。光照时间对生根和茎叶的影响,指出这些参数要进一步研究。
Raiasekaren与Mullins(1979)指出4℃冰冻预处理胚72h增加了产生胚的花药。相反,同样处理30天,会抑制胚的产生。后者归之于相关的诸控制机理的改变。从体细胞胚产生胚的最适温度范围是25-35℃。可是,较高温度对诱导产生的胚质和生活率有不利影响,由于许多褐化和产生类愈伤组织生长。温度低于15℃,抑制了这种胚的产生。
(2)竞争性表达 以胡萝卜典型系统言,PEM细胞转变成胚,产生于减少生长素的培养基中。矿质离子( 和K+)加强这种转变,除生长素外,其它生长调节剂则产生抑制作用。当培养体中非PEM细胞(大,高度液泡化细胞)占优势时,竞争性消失。这是由于非PEM细胞向细胞培养物外伸或PEM转变成非PEM细胞或二者尚待研究确定。
和K+)加强这种转变,除生长素外,其它生长调节剂则产生抑制作用。当培养体中非PEM细胞(大,高度液泡化细胞)占优势时,竞争性消失。这是由于非PEM细胞向细胞培养物外伸或PEM转变成非PEM细胞或二者尚待研究确定。
葡萄花药愈伤组织转变成可识别的胚前体,也需要培养在无荷尔蒙培养基上,与前述胡萝卜情况相同。相反,胚珠和营养组织产生的愈伤组织,显然要求从一种较活跃生长素转换成较不活跃生长素。这种转换:胚珠愈伤组织是2,4-D→NOA→无荷尔蒙培养基,营养组织的是2,4-D→NAA→无荷尔蒙培养基,才能产生有生活力胚。体细胞胚表皮产生的体细胞胚的竞争性的表达是自发的。基于用细胞分裂素或低温处理的胚表面,未能观察到胚柄的事实,作出这种结论。
采用葡萄PEM细胞簇为原生质体源,比用其它组织或细胞较好,因为后者(至少葡萄的)尚未能产生器官建成竞争性。
(3)胚发育 葡萄种子需低温5℃60-90天才能发芽。花药,胚珠,叶柄和体细胞胚产生的胚,发育慢,常不能产生正常植株。花药产生的胚,发育成绿色子叶,但不能形成茎叶,体细胞胚产生的组织不能形成分开的、扩张的、绿色子叶和茎叶。花药产生的胚经4℃2-8周后发育正常,而胚珠发育成正常植株,要求培养在加有细胞分裂素-赤霉酸培养基,继之无荷尔蒙的盐用量降低的培养基。试用几种物理和化学因素,从次生胚产生正常再生植株,至今未能取得满意结果。冷冻、细胞分裂素、赤霉酸、细胞分裂素+赤霉酸,不加蔗糖继之以再饲喂,对正常植株发育取得进展(绿色化和子叶分开),但至今没有一种因素能增高正常植株产生频率(5-30%)。
从体细胞胚表皮启动体细胞胚,显然受到一种相关控制。发育正常的胚极少产生体细胞胚。相反,带有休眠子叶和芽分生组织的胚,产生胚频率近于80%。观察到产生正常植株和次生胚产生胚频率的季节变异。冬季前者可达30%,胚产生频率几乎是零。相反,夏季则反之。促进正常植株发育的理、化因素,抑制体细胞胚胎发生。植物发育与体细胞胚形成间的相背关系可为研究休眠和胚胎发生机理提供有价值手段,或将揭开植物发育一套新的相关控制机理。
(4)再生植株的遗传稳定性和表现葡萄Seyval愈伤组织再生植株的几种特性与亲本不同。例如,叶深蓝绿,幼茎红色而不是通常绿色,和果簇圆筒而不是圆锥形。值得注意这些现象与原来的杂种的相一致。3年生再生蔓在20Brix每英亩产葡萄5t。再生植株切株易生根,比亲本表现一致和地上部生长旺盛。设想这是由于消除了亲本的潜伏病毒的结果。
5.微繁殖 葡萄茎尖繁殖有二种方法。一是传统方法,从茎尖或侧芽取小外植体,诱导产生复生侧芽,继而生根。另一是从茎尖切段切取小叶,诱导产生不定芽。
(1)外植体源一般从冷冻处理的木质茎取快速扩伸芽的1-2cm顶端外植体,或在快速生长时期从大田取样。
(2)消毒 用次氯酸钠(1.2%w/v)或钙(5%w/v)加吞温-20(0.01-0.1%v/v)浸外植体10-20min,无菌蒸馏水淋洗3次,每次5min,或浸入70%乙醇1min,无菌水淋洗。从无菌外植体上切取0.5-4mm茎尖切段,或将小尖帽盖解剖成约20切段,接种在芽诱导培养基上。含有少于2个叶原基的切段,不能产生不定芽尖。
(3)芽诱导培养基 MS或Jona与Webb的盐类足以芽诱导。维生素和有机成分,生长调节剂除外,除一种例外外,余同MS有机物组成。这种情况中,采用Galzy(1977)维生素成分,减去泛酸钙和生物素。
全部研究采用5-20μM细胞分裂素和0-0.5μM生长素,满足了芽的发育。BA是最有效。一般言之,芽诱导并不需要生长素,但Chee与Pool(1982)发现葡萄Rougeon培养物的最适芽形成需要0.5μM NAA。诱导培养基附加0.3mM ADE和磷酸氢钠(1.4mM)增高芽的增殖。
(4)根启动 葡萄茎尖生根不难。将1.5-3cm芽插入琼脂培养基,或放在液体培养基中的滤纸条上。有些栽培品种生根不要生长素。可是,培养基附加0.1-2μM IAA,NAA或IBA,对稍难生根的是需要的。盐浓度对根启动无影响,当生根培养基盐含量降低时,根生长有所增强。在可见第一个根原基后,从根启动培养基中取去插技,根生长也会增强。
为了适应温室和大田条件,将带根插枝移入Jiffy7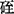 石钵,加玻璃罩,生长9天。除去罩,2周后移入装有堆肥∶沙土(1∶1v/v)混合物的钵中,移置于温室。另一方法,将插枝直接移入钵中,或潮湿繁殖苗床上,或其它保湿方法下,直接生根。
石钵,加玻璃罩,生长9天。除去罩,2周后移入装有堆肥∶沙土(1∶1v/v)混合物的钵中,移置于温室。另一方法,将插枝直接移入钵中,或潮湿繁殖苗床上,或其它保湿方法下,直接生根。
(5)再生植株的遗传稳定性和表现常规茎尖和茎尖切段法,一年内单个茎尖能产生20-24,000植株。
分段的茎尖再生植株含染色体2n=38。可是,它们出现幼年特征(完全叶、无卷须,和轮生叶序),当从人工培养条件下取出植株,它们消失了。由于茎尖切段产生的是不定芽,其遗传变异的或然率,比用标准方法使固有芽发育而成的相对较多。
Harris与Stevenson(1979)建立了包括葡萄×葡萄属杂种在内的14栽培品种的茎尖培养体。其中根砧木5BB,5C和SO4和Vinifera,Pinot Noir,Okanagan Rieshing,和Rotberger难于建成。Barlass与Skene(1980)测试了1l栽培品种,发现砂地葡萄(Rupestrisst.George)和砂地葡萄×冬葡萄(R-99)增殖速率,不象葡萄5个,V。champini2个栽培品种和复合杂种根砧木Harmong的那样快速。
茎尖繁殖或叶切段上诱导不定芽产生的植株,显然有商品潜在价值。可是,需要进行大田观察决定。第二要计算二种方法繁殖所需成本。
6.消除病原菌 近于全部葡萄都是无性法繁殖的。有些克隆可追溯到罗马时代或更早。古老栽培品种的连续克隆繁殖,不经有性循环,有利于病毒,类菌质体生物体和真菌和病菌病原菌的侵染,繁殖和传播。
消除病原菌方法包括全株或离体茎尖,核仁胚胎发生体的热处理,和热处理的茎尖顶帽。
离体茎尖热处理或从热处理植株的茎尖的栽培品种用于消除病毒,有二种问题阻碍着立即应用。最重要问题是热处理不能消除全部病毒,第二难于调节离体,非维管束的茎尖分生组织生长与发育。
(1)茎尖微嫁接 微嫁接是涉及上述问题的最新解决方法。用热处理植株的离体茎尖顶帽嫁接在适宜根砧木上。将带有2-5叶原基长0.1-0.5mm芽分生组织,放在去头幼苗的维管束环上,或插入立体凹陷切口,取自无菌、绿色,根砧木切段。嫁接后3周发育成芽。
嫁接结合的成功,有赖于接穗大小。茎尖顶帽愈大,结合成功率愈高。当顶帽0.5mm或更大时,结合成功率高于50%。可惜,随着顶帽增大病原菌繁殖几率愈高。
采用幼苗根砧木,由于其嫁接结合生存率变异大,并不合宜。从根砧木种取体细胞胚,可为嫁接的顶帽提供遗传一致性更高的受体。曾提出用这种方法消除热稳定病毒。
(2)茎尖培养 从离体的、热处理的茎尖顶帽产生接穗或根砧木,采用人工培养和继而嫁接,为产生无病毒植株提供技术上简便方法。可是,也有困难。上述茎尖外植体的生长与其大小成比例。小顶帽0.2mm或更小,不易伸长,一旦伸长还可能难于生根。目前,许多研究着眼于理、化因素,它们改变茎尖顶帽(2-4叶原基)的生根反应,和在较低程度上人工培养顶帽的生长。所以着重于根启动而不是芽伸长,可能与这种事实有关,就是一旦生了根,这种小外植体将可从这些根取得芽生长因子的供给。无根的芽伸长慢,或不伸长。可是,无根的侧芽和小茎尖外植体能在加细胞分裂素培养基生长,提出可能包含一种新的热处理方式。
(3)根启动和生长 小茎尖外植体上根的启动,是生长素浓度和类型和葡萄基因型的函数。IAA或IBA(0.1μM)时,葡萄外植体生根百分较好,NAA则稍差(Grenan,1979)。相反,砂地葡萄外植体用NAA(0.1μM)生根百分比吲哚类生长素的为高。生长素浓度高于0.1μM,引起产生基部愈伤组织,并降低茎伸展。生长素处理时间和浓度,影响着根伸长的启动和继后生长率。在无荷尔蒙培养基上,根伸长较快。
赤霉酸抑制根形成,但促进其伸长。其它生长调节剂对根启动和生长的影响,未见报道。
在knop盐类(其中缺 )中加K+和
)中加K+和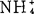 或单加
或单加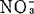 ,增高砂地葡萄外植体的生根百分数。K∶
,增高砂地葡萄外植体的生根百分数。K∶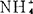 比率高,有利于根启动和生长,但一经启动,降低
比率高,有利于根启动和生长,但一经启动,降低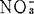 或
或 浓度有利根的长期生长。对生长素或增高K+∶
浓度有利根的长期生长。对生长素或增高K+∶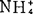 浓度生根反应大致相等,将二者配合,并不能增高生根。外植体培养在加赤霉酸和K+∶
浓度生根反应大致相等,将二者配合,并不能增高生根。外植体培养在加赤霉酸和K+∶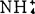 培养基上,根生长被抑制,即使每种处理单用时,能促进根伸长。
培养基上,根生长被抑制,即使每种处理单用时,能促进根伸长。
蔗糖或蔗糖与六种生长素相配合,对根启动无巨大影响。可是,长时期的根生长,增高蔗糖和维生素浓度,有所促进。
(4)芽(茎叶)生长 带有扇叶病毒征状的蔓尖外植体生根百分低。热处理离体芽(35℃90天)生根百分增高,取得无病毒征状植株。把病毒侵染的茎尖外植株经35℃处理5天,比未经侵染的切枝生根百分增多,但未道及病毒征状的消除。
如果启动了根,小茎尖外植体上芽启动发育加快,但无根时,芽生长慢或停止。因此,加强根启动和生长的许多因素,也有利于芽发育。例如,生长素和增高K+, ,
, ,蔗糖,维生素水平和热处理都能增强芽发育。推迟根发育的诸因子,也阻滞芽发育,例如,生长素长期存在或增高矿质盐类。
,蔗糖,维生素水平和热处理都能增强芽发育。推迟根发育的诸因子,也阻滞芽发育,例如,生长素长期存在或增高矿质盐类。
热处理和长期人工培养,能在再生植株中产生表型变异。如曾观察到叶形态改变,花簇数和大小受抑制,在正常光叶上产生毛茸,茎红色素加深,和产生幼年状征状(缺少卷须,叶序改变和叶形)。有些变异是短暂的,只有植株保持在人工培养下才有出现,而其它则十分稳定。
总之,热处理法取得成功,部分有赖于小茎尖切段的生存和发育。很小外植体的生存,为消除热处理稳定病毒所需,可在适宜的盐类和有机组成培养基中加细胞分裂素,可能达到。
珠心胚胎发生性产生无病毒克隆的可能性,应采用病毒侵染的和未经侵染的蔓产生珠心胚。热处理诱导产生的巨大表型变化,可为了解与阶段改变相伴发生的基因表达调节提供新方法。
7.胚和胚珠培养 无籽葡萄的几个栽培品种产生胚,在开花后不久败育。拯救无籽栽培品种的胚,可得无籽特性的有性传递,和其遗传分析。
最近Cain指出无籽栽培品种胚珠培养产生了种子。经冷冻层积后种子发芽。从中取得的胚不能发芽,当培养在含有细胞分裂素盐类培养基上,产生了植株。
Galzy与Galzy(1964)指出从自花授粉的葡萄Aramon种子取得的胚,其发芽需有椰乳(10%v/v)。待根长超过1cm后,胚不再需椰乳。从扇叶病毒侵染的Anamon种子取得的胚,生活力低,许多种子无胚或含有死胚。
有25%种子带有温度敏感基因,调节着叶绿体发育。当将“突变体”幼苗生长在20℃,出现花叶。可是,当花叶植株生长在35℃时,全是绿叶。在叶形成期间,改变温度可操纵相继发生的叶是花叶还是绿叶。
8.调节代谢产物的生产 研究葡萄代谢产物的生产,目的在于了解有机酸的分配,和测验花青素的生物发生。
采用离体花簇人工培养,以测定未成熟葡萄浆果是否积存或合成苹果酸和酒石酸。发育慢的浆果含有苹果酸盐但无酒石酸盐,指出后者可能是从其它来源输入的。
不要荷尔蒙葡萄愈伤组织的花青素生产,因蔗糖浓度增高,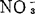 浓度减低,和蔗糖与光的各种组合而增多。并与细胞生长成反相关。由于在蔗糖低浓度中加渗透活性浓度的NaCl,色素生产可得加强,可知蔗糖增加色素生产,是由于培养基渗透性潜势增大所致。
浓度减低,和蔗糖与光的各种组合而增多。并与细胞生长成反相关。由于在蔗糖低浓度中加渗透活性浓度的NaCl,色素生产可得加强,可知蔗糖增加色素生产,是由于培养基渗透性潜势增大所致。
9.逆境生理 测验了NaCl对几种葡萄和杂种栽培品种的茎尖切段诱导产生的不定芽和建成的茎尖培养体的影响。5mM NaCl增强芽的增殖,50mMNaCl有抑制作用。茎尖延长生长在含0-100mMNaCl培养基上继代11次的证实了诸栽培品种对NaCl敏感性不同。在富含NaCl培养基上,V.vinifera栽培品种生长旺势较高于杂种,由于杂种在无盐下生长较差,所以这种差异,不能完全归之于NaCl。Cabernet Sauvignon和Ramsey在富含NaCl培养基上经几次继代后,恢复了生长率。后者在50mM NaCl下仍能恢复,前者恢复NaCl浓度为20μM。比较茎尖培养体与带根插枝的耐NaCl性,提出耐性最高和最低的栽培品种间相当一致。采用芽培养体作为在盐碱条件下根砧木生长的初级筛选显然是可行的。培养体同化高浓度NaCl的可能性尚待研究。根砧木茎尖培养体对供试逆境的适应,可能对葡萄改良是一种有价值手段,由于只要植物是用无性繁殖产生的,这类适应性仍属稳定。
10.离体培养下蔓的发育 葡萄幼年状态可经芽外植体的反复继代或诱导不定芽或体细胞胚诱导产生。培养基内加细胞分裂素,经增高不定芽的产生或早期侧生分生组织发育加速幼年状态的出现。培养基附加诸维生素或CH能推迟人工培养芽切段转入幼年状态。有些栽培品种在加维生素时产生卷须,有的产生卷须、花和果。当把人工培养生长植株移到大田条件时,立即转变成成熟型。
幼年状态的可用于葡萄改良的一个有价值特性,是易于器官回复到幼年态生长。许多木本多年生植物的幼年型易于生根,而成年型则不能。其幼年组织的愈伤组织或组织外植体再生不定芽或胚,而从成年组织产生的愈伤组织可能仍然无变化(Mott,1978)。所以,用葡萄或其它木本植物的茎尖外植体经反复继代诱导幼年性,可供产生带有胚胎发生竞争性的细胞,或具有较大再生潜力的原生质体,提供有益的起点。这种方法优点是幼年组织的克隆基因型能以保藏。
葡萄幼苗须经3-5年才能进入成年或开花时期。缩短幼年期可加速葡萄育种。
葡萄卷须和花是同源器官,起源于一组独立的分生组织细胞,叫原基或化学长链,后者决定着哪种器官将产生原基。细胞分裂素诱导产生的高度分枝的原基,产生花原基,而GA3诱导产生的弱分枝原基产生卷须。用细胞分裂素处理6月龄蔓的卷须,导致花和果形成。用各种细胞分裂素人工培养的卷须,形成退化花,无雌和雄孢子体。人工培养的花有丝分裂细胞内调节减数分裂的诸因子尚待鉴别。
细胞分裂素处理完整植株,产生可结实的可育花,而人工培养的卷须产生的花,而雌、雄孢子体。后者不发生减数分裂,可能由于木质部和韧皮部内某种易位的因素。每种组织系统的渗出物,可为完全花发育提供所需的诸因子。鉴定“减数分裂”调节物,有益于人工培养花的产生,并可为从二倍体产生单倍体细胞提供方法。
离体培养外植体对平衡的生长调节剂的反应,使独立分生组织转变成花器状态,为探索花器的诱导的基因调节机理及其联合的代谢提供有价值手段。
11.寄主-寄生物互作 Morel(1948)证实了Plasmophora viticola Berl.和Toni(粉霉病)在葡萄游离上连续繁殖。寄主和病原菌的共存,为阐明下列问题提供机会:(1)细胞培养能用来筛选耐粉霉病克隆吗?(2)活体和人工培养中寄主与寄生物互作是否一致?(3)寄生物生长是否需要寄主的特殊代谢产物?正在发掘有关这类问题的体系。
浓杆菌部分质粒DNA转给寄主(双子叶植物)导致形成肿瘤(Chilton等,1977)。这种细菌的寄主范围,受质粒基因的控制(Thomashow等,1980)。已鉴别出质粒的二个基础群;侵染大量属和具有广谱寄主范围(WHR)质粒,侵染原始葡萄和偶及树莓和具有有限寄主范围(LHR)质粒。WHR质粒的T-DNA(诱导肿瘤DNA)与LHR质粒的DNA只有极少数共同顺序,指出启动肿瘤的基因是不同的。
含有WHR质粒的浓杆菌,不能在葡萄上产生肿瘤,但Krul等在葡萄Seyval上发现,由细菌的WHR与LHR菌系引起形成肿瘤。再者,作者证实了WHR和LHM菌系的DNA,渗入了Seyval DNA中。
在采用这种质粒作为葡萄遗传转化的穿梭载体前,需要肯定WHR和LHR质粒DNA顺序相一致,和它们与葡萄寄主范围的关系。具有非同源DNA顺序使葡萄产生肿瘤,可能深入了解诱导肿瘤的遗传基础。
(三)展望
Meredith(1981)认为在材料和人力充裕条件下,用基因载体和针对理、化逆境下选择变异体,潜在葡萄遗传改良成功的或然率与大多数其它属的相同。
障碍是可克服的。需要植株再生潜力高的原生质体系统。器官建成竞争性的原生质体的可能来源,可能是体细胞胚,或经反复继代导致产生幼年特征的茎尖。同样,大多数葡萄栽培品种缺乏人工培养器官建成潜势的情况,可采用从诱导产生的幼芽作外植体予以改善。
浓杆菌的LHR T-DNA的分离和鉴别和更一步阐明寄主-质粒互作,将有利于建立用于葡萄的基因载体。克服上述障碍后,就要决定什么基因或基因小群能增高葡萄生产力。采用缩短蔓的幼年时期的新方法,有希望取得成就。
【参考文献】:
〔1〕Arora,I.K.and R.N.Singh 1978 Growth hormones and in vitro callus formation of papaya.Sci.Hortic.8∶357-381.
〔2〕DeBruijne,E.,E.DeLanghe,and R.van Rijck 1974 Action of hormones and embryoid formation in callus cultures of Carica papaya.Int.Symp。Fytofarm.Fytiat。26∶637-645.
〔3〕Litz,R。E,and R.A.Conover 1978 In vitro propagation of papaya.Hortscience13∶241-242.
〔4〕Litz,R。E。1981 In vitro polyembryony in Carica papaya L。ovules.Z.pflanzen-physiol.104∶285-288.
〔5〕Yiu,S.an.d S.I.Liaw 1977 Plant regeneration from shoot tips and callus of papaya。In vitro 13∶564-567.〔6〕Krul,W.R.and J.Myerson 1980 In vitro propagation of grape.In∶Proc.Conf.on Nursery Production of Fruit Plants through Tissue Culture.Applications and Feasibility(R.Zimmerman,ed.)pp.35-43,USDA Sci.and Education Adm-inistrotion,Agric.Res.Results,ARR-NE-11,Beltsville,Margland.
〔7〕Lai,P.C.and H.T。I,1980 Studies on tissue culture of grape(Vitis viniferaL.):Influence of environmental factors on the growth and differentiat ion of gra pe callus.J.Agric.Res.China,29(2)∶157-165.
〔8〕Meredith,C.P.1981 Genetic engineering-The outlook for grapes,Wine Inves-tor 5∶1——4。
〔9〕Mowbray,G.H.1981 Clone me around again,Willie.Virginia Farm Vine-to-Wine letter 1∶4-5.
〔10〕Olmo,H.P.1976 Grapes.In∶Evolution of Crop Plants(N.W.Simmonds,ed.)pp。294——298,Acad.Press,New York.
〔11〕Hadaka,T。,Y.Yarnada and T.Shichijo 1979 In vitro differentiation of haploid plants by anther culture in Poncirus trifoliata(L.)Raf.Jpn. J.Breed.29∶248-254.
〔12〕Maheshwari,P.and N.S.Rangaswamy 1958 Polyembryony and in vitro Culture of embrvos of Citrus and mangifera.Zndian J.Hortic。15∶275-282.
〔13〕Navarro,L。,C.N.Roistacher and T.murashige 1975 Improvement of shoot-tip grafting in vilro for vitus free clones.J.Am。Soc.Hortic.Sci.100∶475-479.
〔14〕Spiegel-Roy,P.and J.Kochba 1980 Embryogenes is in Citrus tissue cultures.In Advances in Biochemical Engineering(A Fi echt er,ed)Vol。16,pp.27-48, Springer Verlag,Berlin,Heidelberg.
〔15〕Vardi,A.1982 Protoplast derived plants from different Citrus Species and cultivars.Proc.I ntern.Soc.Citicultune.(1981)1∶149-152.