植物对养分的吸收
出处:按学科分类—工业技术 中国农业出版社《肥料实用手册》第4页(7794字)
【参考文献】:
[1]何念祖,孟赐福.植物营养原理.上海:上海科学技术出版社,1987
[2]张道勇,王鹤平.中国实用肥料学.上海:上海科学技术出版社,1997
[3]浙江农业大学.作物营养与施肥北京:农业出版社,1990
[4]谭金芳,介晓磊等.钾肥施用原理与实践.北京:中国农业科技出版社,1996
[5]林葆.我国化肥的肥效及其提高的途径.土壤学报.1989
[6]沈其荣,谭金芳,钱晓青.土壤肥料学通论.北京:高等教育出版社,2001
[7]陈伦寿,李仁岗.农田施肥原理与实践.北京:农业出版社,1984
[8]浙江农业大学.植物营养与肥料.北京:农业出版社,1988
植物根系是吸收养分的主要器官。根系吸收的养分被运到植物的地上部分,并且在植物体内进行再分配。无机离子的吸收——跨膜运输是一个重要的生理过程,它不仅决定了离子在细胞中的积累,而且影响着植物的生长发育。
(一)植物对养分吸收的特性
1.选择性 植物从介质中吸收无机离子,并不决定于介质中离子的浓度,而是由植物根系自身的需要优先吸收某些营养元素,而摒弃另一些元素。例如,海水或池水中钠的浓度大大高于钾,虽钠和钾的化学性质很相似,但植物却能选择性地吸收更多的钾。
2.基因型差异 不同植物或同一植物的不同品种,吸收与运输养分的能力有所不同,这是由于在植物中存在着差异性所致。例如,用相同培养液分别培养番茄和水稻,番茄对钙和镁的吸收速率快,吸收量多,但几乎不吸收硅。水稻对钙和镁的吸收速率较慢,吸收量也少,但却大量地吸收硅。近年来这方面的研究极为活跃,人们希望通过育种的手段,筛选或培养吸收养分能力强的品种,以适应在贫瘠土壤上生长,或选育利用肥料养分能力强的品种,以提高肥料的利用率。
3.累积性 植物在吸收无机离子时,尽管植物细胞中的离子浓度比外界溶液浓度高得多,但仍能吸收无机离子。海带中I-浓度远高于海水中I-浓度,但仍能吸收大量的碘。在通常情况下,这种吸收是逆浓度、逆电位势进行的,一般需要能量的供给。
4.阶段性 植物营养期包含若干营养阶段,不同的阶段对营养元素的种类、数量和比例即营养条件有不同的要求,这就是植物营养的阶段性。比如,水稻在苗期吸收N、P2O5、K2O的百分数分别为0.50%、0.26%、0.40%;分蘖期吸收三者的百分数分别为23.16%、10.58%、16.95%;圆秆期吸收量则增加为52.40%、58.03%、59.74%。
(二)植物对养分吸收的形式
1.介质养分向根迁移的方式 养分离子向根部的迁移,一般有截获、扩散和集流3条途径(图1-1)。

图1-1 养分离子向根部迁移一般通过3种途径
①截获 ②集流 ③扩散
(1)截获:根系从土壤中吸收养分时,由于植物根与土粒密切接触,因此,根细胞外H+和黏土外交换性阳离子发生离子交换。这种由于接触土壤而获得养分的过程叫截获。通过截获吸收离子态养分极少,但钙离子经截获移至根部一般较多些,其次是镁离子。
(2)集流:由于植物的蒸腾作用,使养分随水流通过土壤孔隙向作物根部的迁移即集流。它主要受植物的蒸腾作用和土壤溶液中离子态养分多少的影响。当土壤温度高时,植物蒸腾作用较大,体内失水较多,使根际周围水分不断地流入根表,土中离子态养分也就随着水流达到根表。当土壤中离子态养分含量较多,供给根表的养分也随着增加。氮、钙、镁主要是由集流供给的,而且钙、镁供应量常能满足一般作物的需要。
(3)扩散:在土壤溶液中当某种养分浓度出现差异时,所引起的养分迁移到根表的过程。这一作用使养分从浓度高处向浓度低处移动,直至达到平衡。当根对养分的吸收大于养分由集流迁移到根表的速率时,根表面养分离子浓度下降,同时根周围土壤中养分也有不同程度的减少,出现根际某些养分亏缺,于是根表与附近土体间产生浓度梯度,高浓度养分向低浓度处扩散,土壤中养分则向根表迁移。植物养分在土壤中的扩散受很多因素影响,如Cl-、K+、NO3-在水中扩散系数较大,而PO43-则较小,土壤质地较轻的砂土,养分扩散速率比质地较重的土壤快,土温高的比土温低的扩散快等等。
应该说以上养分迁移的3种过程在土壤中是同时存在的。但总体来说,截获取决于根表与土壤黏粒接触面积的大小,集流取决于根表与其周围水势的大小,扩散取决于根表与其周围养分浓度梯度的高低,它们都与根系活力有密切关系。
2.植物对养分的吸收机制 土壤溶液中养分离子通过集流与扩散以及部分根部截获到达根表面,吸附在细胞表面。这些离子先进入自由空间,即从细胞壁到原生质膜的空隙,还包括细胞间隙,然后进入根系的内皮层。
(1)被动吸收:是顺着电化学势的梯度运输,即离子在细胞的自由空间内,可以通过扩散或质流经细胞膜向膜内移动;同时是不需要代谢供应能量的吸收作用。包括以下两种:
A.简单扩散:当细胞外溶液的浓度大于细胞内时,溶质分子就会由于扩散作用进入细胞内。
B.杜南平衡:植物在吸收营养物质时,常有这样的情况,细胞内某种离子的浓度已经超过了外界该离子的浓度,该离子仍可逆浓度梯度扩散而进入细胞内,达到平衡。这种现象只有用杜南(F.G.Donnan)提出的平衡理论才能说明。
杜南指出,细胞内有不能扩散出的阴离子(R-)(如蛋白质离子)存在,它在细胞内必然会吸引一些阳离子如Na+,而以盐的形式存在于细胞内,使细胞呈电中性。将这个细胞放入NaCl溶液中,因为细胞内没有Cl-,细胞外的Cl-势必向细胞内扩散,由于电荷不平衡,Na+也跟随而进入,当细胞内可扩散正、负离子浓度的乘积和细胞外可扩散的正、负离子浓度的乘积相等时,达到平衡状态。这种平衡称为杜南平衡,其表达式为:

植物细胞通过杜南平衡使细胞积累离子的过程,并不需要代谢能量做功,是个被动吸收离子的过程。
大多数阳离子(除K+外)都是属被动吸收。
(2)主动吸收:是逆电化学势梯度的运输,需要能量,所有的生活细胞,与外部介质相比较总是带负电荷,而内部阴离子浓度绝对高于平衡浓度,所以带负电荷的阴离子如NO3-、H2PO4-、SO42-、Cl-是逆电化学势梯度的吸收,属于主动吸收。目前,从能量的观点和酶动力学原理来研究植物的主动吸收,提出了载体学说和离子泵解说。
①载体学说:此学说是以酶动力学说为其理论依据。用此原理将载体与养分的关系比作酶和底物的关系:

应用米凯利斯—门藤(Michaelis—Menten)方程式,可求出酶促反应的速率(v),最大速率(V)和米氏常数(Km)。

当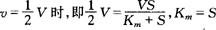
当酶促反应速度等于最大速度的一半时,此时的底物浓度就是Km。由于
它与k1成反比。因此,Km值愈小,表示酶对底物的亲和力愈大,酶促反应速度则愈快。所以,Km值是酶对底物亲和力的倒数。
依据上述原理,对各种作物根或叶片在不同浓度溶液中吸收各种离子的速度进行研究,可求出Km值(表1-2)。例如观察甘蔗离体叶片在30℃时吸收Zn2+和Cu2+的速度,测定其Km值分别为0.0111mmol/L和0.0145mmol/L,表明甘蔗叶片对Zn2+和Cu2+的亲和力基本相当;而将大麦离子根放在不同浓度氯化钾溶液中,观察30℃吸收K+的速度,测定其Km值为0.021mmol/L,Na+则为0.32mmol/L,表明大麦根对K+的亲和力远大于Na+。所以当溶液中K+和Na+同时存在时,则可选择吸收K+。这与各种酶对于不同底物具有不同的Km值很相似。
表1-2 各种作物对各种离子在低浓度下的米氏常数(Km)

载体究竟是什么化合物,至今不太清楚。一般认为载体是生物膜上含有一定的能载运离子越过膜的分子。载体分子对一定的离子有专一的结合部位,能选择性地携带某种离子通过膜。载体的活化需要ATP,其转运离子的情况见图1-2。在膜的外部界面上,活化载体遇到与它具有亲和力的离子结合成载体——离子复合物,转移到膜内,与磷酸酶接触,能裂解出磷酸基,使载体失活,因而离子被释放到膜内的细胞质中,失活的载体在膜中磷酸激酶和ATP的作用下又可使其磷酸化,而再度活化,转移到膜朝外面的一边,再与膜外离子形成复合物。如此重复循环,就把膜外的物质不断运入膜内,积累于细胞内。载体磷酸化所需的ATP可由根的呼吸作用提供。

图1-2
A.离子通过膜的传递模式 B.配合能量消耗
(Marschner,H.,1986)
主动吸收过程有载体存在,可以从下列两方面得到证明:
第一,饱和效应。在研究离子吸收与外界离子浓度的关系时发现,细胞对离子的吸收有个最大的极限值。在外界离子浓度比较低的范围内吸收速率与离子浓度成正比,当离子浓度超过某个数值以后,随着外界离子浓度的增加,吸收速度的增加会变得缓慢,最后细胞对离子的吸收达到饱和后,就不再吸收。这种现象称为饱和效应,即一种特定的离子有一种特定的载体,当载体上的结合部位被饱和后,尽管外界溶液中离子浓度增加,主动吸收的速率也不再增加。
第二,离子竞争。研究不同的离子在吸收过程中的相互关系时发现,离子吸收表现竞争性的抑制。例如,将大麦根浸在含Cl-的溶液中,再加入Br或I-后,Cl-的吸收就减少,这说明Cl-和Br-或I-可以和同一载体相结合,表现出离子竞争现象。而磷酸或硝酸与氯结合的载体不同,则无竞争抑制作用。
近年来,从酵母和其他微生物的质膜中,已分离出能运载某种分子或离子的透过酶,这就是载体,可是在高等植物中尚未发现。但有人用抗生物质作为离子载体进行试验,发现燕麦培养液中加入短杆菌肽后,燕麦可选择吸收K+,当加入氰酸盐被抑制后,吸收也受到抑制。目前研究离子载体的抗生物质有以下几种,见表1-3。
表1-3 离子载体(抗生物质)的特性

离子载体如缬氨霉素是一种环状多肽,环外是非极性的,能溶于膜脂,环内有羰基,能与K+结合形成复合物,使K+能越过膜的非极性区,进入细胞内。由于离子载体对各种离子有选择性,所以植物就有选择性的吸收。
总之,载体学说认为,物质透过膜是由于膜中的载体和物质结合后,在膜内从一面转向另一面(即转动180°),再将该物质释放出来。可是膜和载体的两侧都是极性的,中部都是非极性的,载体在膜中是难以转动的。因此,转运机制尚无定论。
②离子泵学说:泵是指能量的意思。离子泵即可以在逆电化学势陡度的情况下将离子泵入或泵出细胞膜,实现主动运输的机理,这种泵能使细胞的离子成分和浓度维持稳定并不同于外界环境,离子泵能够在环境中溶质含量非常低的情况下吸收和浓集离子,以致使细胞内外离子含量有很大的不同。
离子泵学说是由Hodges(1994)所提出,该学说认为,质膜内的ATP酶能将ATP水解为ADP和Pi,释放出的能量可将离子泵入细胞内,如图1-3所示。图1-3说明了植物细胞吸收阳离子和阴离子,也描绘了ATP酶和阴离子载体。由于ATP酶的作用,使ATP水解,产生阴离子ADP-和阳离子H+(ATP+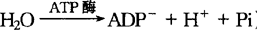 )。H+释放到膜外,ADP留在膜内。因而产生跨膜的pH梯度及电化学势能,使膜内有更多的负电荷,膜外的阳离子被负电荷吸收进入细胞内。另一方面,使ADP-与H2O反应,增加了细胞内的OH-浓度。
)。H+释放到膜外,ADP留在膜内。因而产生跨膜的pH梯度及电化学势能,使膜内有更多的负电荷,膜外的阳离子被负电荷吸收进入细胞内。另一方面,使ADP-与H2O反应,增加了细胞内的OH-浓度。
ADP-+H2O→ADP+OH

图1-3 ATP酶启动阳离子泵并与阴离子载体相偶联
细胞质中的OH-能启动阴离子载体,OH-被排出,使其他阴离子(如SO42-)被细胞吸收,离子吸收所需的能量由ATP酶催化ATP水解提供。
(3)叶部吸收(根外营养):植物不仅依靠根部从土壤中吸取养分,还能通过叶部等地上部分吸收养分,这一现象称为根外营养。把肥料配成一定浓度的溶液喷洒在茎叶上叫根外追肥。
叶部吸收养分一般是从叶片角质层和气孔进入,最后通过质膜而进入细胞内。一些试验表明,叶部吸收养分的机构与根部吸收一样,如玉米叶片吸收K+的Km平均值为0.038mmol/L。-般来说,在植物整个营养生长期间,叶部都有吸收养分的可能,但是吸收的强度则不同。因为植物吸收养分不仅取决于养分的种类、浓度、介质反应、溶液与液面的接触时间以及植物吸收器官的年龄,而且和植物体内的代谢作用关系更大。
植物根外营养的特点主要表现在以下几方面:第一,直接使作物吸收养分,避免养分在土壤中固定。如在石灰性土壤或盐渍土中,铁多呈不溶性的3价态,植物难以吸收,常患缺绿症。在红黄壤上栽培果树,常发生微量元素不足,如浙江省黄岩山地黄壤桂圆土壤缺锌,北方石灰性土壤缺铁也很普遍,根外追肥会使肥料效果显着提高。第二,肥效快。吸收运转养分的速度比根部快,能及时满足作物需要。例如,豆科植物生长后期喷施少量硼肥,可提高种子收成。因为硼能促进花粉萌发,保证花粉管迅速地进入子房,从而保证了种子的形成。第三,直接参与植物体内代谢,从而减缓根系衰老。如甘薯、马铃薯,淀粉在其体内要不断地合成,这属于碳水化合物代谢,而磷、钾与该代谢有着密切的关系。此外,蔗糖在植物体内是以蔗糖磷酸酯作为碳水化合物运输的主要形式,由叶进到块根或块茎中,所以磷、钾肥在淀粉累积中尤为重要。第四,施肥均匀。有些元素作物需量少,土壤施用每667m2也仅需1~2kg,因此,土壤施用难以施得均匀,很可能出现有些地方用量不够不见效,有些地方施得过量而产生毒害作用。而根外追肥可弥补这一不足。第五,经济效益高。根外追肥用量仅是土壤施肥的1/10~1/5,且省工省时,操作简单。农业实践早已证明,栽培块根块茎类作物采用磷、钾根外追肥,不仅能提高产量,而且可改善品质。
根外追肥的效果受到多种因素的影响。首先,各种作物叶面气孔多少不一,角质层厚薄不等,因而根外追肥的效果表现得也有差别。一般来说,双子叶植物如棉花、豆类、油菜、马铃薯等叶面较大,角质层较薄,溶液易渗入叶内,故根外追肥效果较好;而单子叶植物如水稻、小麦叶面较小,角质层较厚,溶液渗透比较困难。因此,在进行根外追肥时,必须混合少量“湿润剂”,如中性皂或洗涤剂,浓度为0.1%~0.2%,使养料易于渗透。其次,各种肥料的透性差别也导致了肥效的不同。如氮肥中的尿素易进入叶面,因此效果较好。如果将尿素和其他矿质肥料混合喷施,也可提高离子态养料的通透速度。第三,调节介质反应,也可使叶部肥效有所改变,如以阴离子为主的(磷酸盐肥料),可调节到酸性,选用过磷酸钙;以阳离子为主的,可以调节到中性至微碱性。试验表明,在酸性反应时,叶部吸收肥料中的阴离子较多,中性至微碱性反应时,吸收阳离子较多。
由于根外追肥是一种用肥省、投资少、见效快的施肥技术,所以在改进施肥技术方面有很大意义。但需指出,根外追肥只能作为特殊情况下的应急措施,不能完全代替根际施肥。
(三)植物的养分吸收量和吸收速度
作物生长期中吸收养分的数量在很大程度上取决于作物种类和产量。作物产量和养分吸收又紧紧依赖于作物栽培品种。由于品种和产量的不同,吸收的养分量差异很大。就品种而言,不同作物的根系长度和表面积差别很大,而作物根系阳离子交换量的大小与根系吸收养分能力强弱有密切的关系。一般来说,根系阳离子交换量大的作物,吸收土壤中阳离子的总量也较多,反之则较少。比如,豆科作物阳离子交换量大,吸收能力强,麦类作物阳离子交换量小,吸收能力弱,而玉米、番茄、马铃薯的阳离子交换量介于其间,吸收力中等。就同一作物来看,根的不同部位阳离子交换量差别也很大,如豌豆根尖的阳离子交换量每100g干根可达125mol,比其根的阳离子交换量平均值高1倍以上。
作物在不同生长时期,吸收的养分量也不等,一般在生长最旺盛时吸收的养分最多,吸收速率最快。因此,在估算作物需肥量时,它的吸收速率是一个重要因素,即一般生长期短,吸收速率快,需要养分多。例如番茄和糖用甜菜每年每单位面积吸收的钾量与甘蔗是相等的,但是,前两种作物生长期只有120d,而甘蔗生长期为1年。这表明吸钾总量相等的作物,生长期短的比生长期长的需钾量要大得多,因为它们的吸钾速率(吸收量/单位时间)相差很大。通常情况下,作物营养生长阶段吸钾量最多,如马铃薯生长前期(只占生长期的1/3)吸收的钾量占总钾量的50%。